
OSWALDO MOOSER’S FOSSILS FROM AGUASCALIENTES, MEXICO: A SYSTEMATIC APPROACH TO LOST AND FOUND SPECIMENS

Palaeontology in the state of Aguascalientes (Mexico) has an extensive history dating back to 1799. However, it was not until the 1950s that Oswaldo Mooser began to formally describe the palaeontological biodiversity of the state, together with other palaeontologists such as Walter W. Dalquest, with whom he even described multiple new species from local discoveries. Much of the fossil material collected by Mooser was donated to different scientific collections, but the location of several specimens was never specified, and they were lost for decades, including holotypes. Bibliographic sources were consulted to trace the location of part of the Mooser’s collection. A total of four local, national, and foreign institutions were consulted to examine the palaeontological material, and a compilation of various fossils collected by Mooser in Aguascalientes was made, resulting in 76 original specimens of which nine are holotypes. Additional bibliographic sources were used to describe and reassess some specimens that had not been properly evaluated before. Previously published information of the fossil material was complemented, and the anatomical description of some specimens is also presented for the first time. Twenty new reports of unpublished fossils collected by Mooser are addressed in this work. The data obtained have implications for the evaluation of the examined taxa, as the specimens documented in this study still require intensive and specific analyses that cover aspects of their systematics, evolution, ecology, and natural history. Thus, the present work aims to serve as a basis for future palaeontological studies.
El Cedazo Local Fauna, Holotypes, Megafauna, Pleistocene, Taxonomy
Figures


![**Figure 6.** The American lion ***Panthera atrox*** [@Leidy1853], right radius bone (**TMM 42428-9793**). **1,** posterior view; **2,** anterior view. Scale bar= 5 cm. **3,** transversal view of the proximal end; **4,** transversal view of the distal end. Scale bar= 2 cm.](https://peapaleontologica.org.ar/index.php/peapa/article/download/533/version/647/945/19047/fig6.jpg)
![**Figure 7.** ***Aguascalientia wilsoni*** [@DalquestMooser1974]. Holotype, **TMM 41536-26**, lateral view. Scale bar= 5 cm.](https://peapaleontologica.org.ar/index.php/peapa/article/download/533/version/647/945/19048/fig7.jpg)



Resumen. LOS FÓSILES DE OSWALDO MOOSER DE AGUASCALIENTES, MÉXICO: UN ENFOQUE SISTEMÁTICO DE EJEMPLARES PERDIDOS Y ENCONTRADOS. La paleontología en el estado de Aguascalientes (México) tiene una amplia historia que se remonta a 1799. Sin embargo, fue hasta la década de 1950 que Oswaldo Mooser comenzó a describir formalmente la biodiversidad paleontológica del estado, junto con otros paleontólogos como Walter W. Dalquest, con quien incluso describió múltiples especies nuevas a partir de descubrimientos locales. Gran parte del material fósil colectado por Mooser fue donado a diferentes colecciones científicas, pero el paradero de varios ejemplares nunca se especificó, y se perdieron durante décadas, incluidos los holotipos. Se consultaron fuentes bibliográficas para rastrear parte de la colección de Mooser. Un total de cuatro instituciones locales, nacionales y extranjeras fueron consultadas para examinar el material paleontológico, y se realizó una recopilación de varios fósiles colectados por Mooser en Aguascalientes, resultando en 76 ejemplares originales de los cuales nueve son holotipos. Se usaron fuentes bibliográficas adicionales para describir y revalorar algunos ejemplares que no habían sido debidamente evaluados con anterioridad. Se complementó la información previamente publicada del material fósil, y también se presenta por primera vez la descripción anatómica de algunos ejemplares. Veinte nuevos reportes de fósiles no publicados colectados por Mooser se abordan en este trabajo. Los datos obtenidos tienen implicaciones para la evaluación de los taxones examinados, ya que los ejemplares documentados en este estudio aún requieren de análisis intensivos y específicos que abarquen aspectos de su sistemática, evolución, ecología, e historia natural. Por lo tanto, el presente trabajo pretende servir como base para futuros estudios paleontológicos.
Palabras clave. - Fauna Local El Cedazo. - Holotipos. - Megafauna. - Pleistoceno. - Taxonomía.
THE FIRST palaeontological discovery in the state of Aguascalientes, Mexico, dates to 1799, when the fossil remains of a member of the order Proboscidea were found in the capital city by Manuel Gutiérrez Solana (Councillor, Perpetual Judge, and Ordinary Mayor of the First Vote of the Villa de Aguascalientes from 1799 to 1808), and published by an anonymous author in La Gaceta de México (Maldonado-Koerdell, 1948; Valencia-Cruz & Guzmán-Gutiérrez, 1994). However, it was not until the decade of 1950 that the state’s palaeontological richness began to be formally described (Guzmán-Gutiérrez & Rodríguez-Ávalos, 2008).
Oswaldo Mooser (Fig. 1) was a Swiss-born chemical bacteriologist who had a great interest in palaeontology, which led him to conduct research in this field and to teach palaeontology to the first generations of the Biology Program students at the Universidad Autónoma de Aguascalientes from 1975 to 1981 (Estrada (1989); María Elena Siqueiros-Delgado and Marcelo Silva-Briano, former students from 1975 to 1980 and from 1976 to 1981, respectively, pers. comm., 27 September 2024). Mooser (1955) published his first contribution to the knowledge of the past biodiversity that inhabited the territory that is now the state of Aguascalientes: an informative publication on the vertebrate Pleistocene fauna of the region, where he mentions some of the palaeontological discoveries made in the area, from small lagomorphs, medium-sized pronghorns, horses, camels, and sabre-toothed cats, to the larger proboscideans (Mooser, 1955).
A few years later, Mooser (1958) described in detail his palaeontological findings in Aguascalientes. The fossil material was referred as “El Cedazo Local Fauna”, since the specimens were recovered in the outcrops of Pleistocene sediments exposed by erosion in the El Cedazo and San Francisco creeks, near the city of Aguascalientes. El Cedazo creek is located eastwards the capital city and extends from the old El Cedazo Dam (now Centro de Educación Ambiental Cultural y Recreativo El Cedazo) to the northeast, in the vicinity of the small-town of San José de la Ordeña, Municipality of Aguascalientes (Tapia-García & Sandoval-Ortega, 2024a). However, Mooser (1958) also documented occurrences of Pleistocene fossils in other localities of the state, like in the Municipality of Calvillo. Eventually, more specimens would be recovered from other municipalities such as Asientos, Jesús María and Rincón de Romos (Tapia-García & Sandoval-Ortega, 2024a).
The taxa reported by Mooser (1955, 1958) correspond mostly to megafauna, animals with a body mass greater than 44 kg (Faith, 2011; Galetti et al., 2018; Johnson, 2002), but smaller mammals were also registered, including a North American porcupine (Hibbard & Mooser, 1963), and even reptiles like tortoises (Mooser, 1972, 1980). Together with the American palaeontologist Walter W. Dalquest, Mooser studied the fossil fauna of the state’s territory and discovered a variety of new species of different lineages.
Dalquest and Mooser (1974) described the Miocene fauna of Aguascalientes and reported four herbivorous mammals of the families Camelidae, Merycoidodontidae, Tayassuidae and Rhinocerotidae (as well as other taxa of vertebrates not discussed, such as tortoises), found at the fossiliferous locality El Zoyatal, which historically was the second known locality with Miocene fauna in Mexico (Dalquest & Mooser, 1974). This locality is currently inaccessible, as it was completely covered after the quarry work from which the fossils were recovered (Guzmán-Gutiérrez & Rodríguez-Ávalos, 2008).
Among the Miocene fauna of Aguascalientes is Aguascalientia wilsoni (Dalquest & Mooser, 1974), a small camelid with an elongated rostrum described as a new species from a fragmentary mandible with preserved teeth (Stevens, 1977). That same year, Dalquest (1974) proposed a new species of Pleistocene antilocaprid: Tetrameryx mooseri Dalquest, 1974, described from cranial remains found in the Arroyo San Francisco locality.
One year later, Mooser and Dalquest (1975a) proposed a new Pleistocene camel species, Camelops traviswhitei Mooser and Dalquest, 1975, from cranial and mandibular remains. Later, the authors reported the presence of at least 39 Pleistocene mammal species in Aguascalientes, including: Canis cedazoensis Mooser and Dalquest, 1975, Tetrameryx tacubayensis Mooser and Dalquest, 1975 and Bison aguascalentensis Mooser and Dalquest, 1975 (Mooser & Dalquest, 1975b).
The latest contribution of Mooser and Dalquest dates to 1980, when additional fossil material of a North American giant short-faced bear from the Middle Pleistocene was found, consisting of a partially complete, well-preserved right jaw with dental material (Dalquest & Mooser, 1980). Previously, Mooser and Dalquest (1975b) had reported the presence of the enormous Arctodus simus (Cope, 1879) in Aguascalientes during the Pleistocene based on a single tooth (currently lost). However, although this species may have occupied the state’s territory, based on the newer available evidence, they concluded that the fossil material from the North American giant short-faced bear found in Aguascalientes corresponds to Arctodus pristinus Leidy, 1854 (Leidy, 1854c) (Dalquest & Mooser, 1980). This fossil is currently deposited in the Museo Regional de Historia de Aguascalientes, along with other fossils from the collection of Oswaldo Mooser (Tapia-García & Sandoval-Ortega, 2024a, 2024b), and among the original specimens collected by him that remained in Aguascalientes, it is one of the best preserved and most complete.
Although much of the palaeontological research in Aguascalientes addressed some of the taxonomical aspects of the extinct megafauna, chronostratigraphic information on fossiliferous localities was limited for a long time (Hernández-Láscares, 1981). Geological studies were made to describe the stratigraphy of the central region of the state (Hernández-Láscares, 1979, 1981). However, it was not until the decade of 1990 that an Irvingtonian (Early to Middle Pleistocene, 1.9 My to 250 ky BP) to Rancholabrean (Late Pleistocene, 250 to 11.7 ky BP) age was determined for El Cedazo Local Fauna, which brought important implications to understand several palaeontological issues (Montellano-Ballesteros, 1990). Furthermore, it was also mentioned that the fossils from the stratum of reddish-brown tuffaceous sand might date to an Irvingtonian age; in contrast, a Rancholabrean age is attributed to fossils that are found in the stratum of whitish fine sandstone. The specimens from both ages tend to develop a similar colouration to that of the stratum in which they are found (Montellano-Ballesteros, 1990). Although the specimen colour may represent a good indicator of the age, the colouration may be relative to the deposit environment and other factors, and the age cannot always be inferred with certainty, especially if there is no additional stratigraphic information of the specimens collected by Mooser (Montellano-Ballesteros, 1990).
All these palaeontological studies made so far in Aguascalientes have substantially contributed to the knowledge of the Miocene and Quaternary (Pleistocene to Present) fauna in North America, especially on mammalian megafauna, since a variety of new species has been described from the fossiliferous localities in the state (Ferrusquía-Villafranca, 1978; Ferrusquía-Villafranca et al., 2010; Kurtén & Anderson, 1980). Therefore, it is pertinent to continue the palaeontological studies to enhance the understanding of the past biodiversity that inhabited North America.
Currently, much of the material collected by Mooser is segregated in multiple scientific collections such as the Colección Zoológica de la Universidad Autónoma de Aguascalientes (CZUAA), Museo Regional de Historia de Aguascalientes (MRHA), Colección Nacional de Paleontología, Museo María del Carmen Perrilliat M., Instituto de Geología, Universidad Nacional Autónoma de México (UNAM) as well as some foreign institutions in Texas, USA (Díaz-Sibaja, 2018; Montellano-Ballesteros, 1990; Reynoso-Rosales & Montellano-Ballesteros, 1994; Stevens, 1977; Tapia-García & Sandoval-Ortega, 2024a, 2024b; Vlachos, 2018) (Stevens, 1977; Montellano-Ballesteros, 1990; Reynoso-Rosales & Montellano-Ballesteros, 1994; Díaz-Sibaja, 2018; Vlachos, 2018; Tapia-García & Sandoval-Ortega, 2024a, 2024b). Allegedly, Mooser donated about 275 fossil specimens from Aguascalientes to the Instituto de Geología, UNAM, in 1959, and approximately 745 to Midwestern State University, Wichita Falls, Texas (Estrada, 1989). However, the location of many of these fossils has changed through the decades, and a considerable number of the material cited by Mooser in the literature remains lost, since there was no mention of where these specimens were deposited and where the rest of Mooser’s collection ended up after his retirement.
The objective of this study is to report the location of several specimens from Mooser’s collection that were considered lost, including holotypes, which have not been studied in detail since their location remained unknown for decades. In addition, this work provides supplementary information to enhance and complement the previous descriptions of the fossil material. Furthermore, this compilation is intended to serve as a basis for future research that will help clarify aspects of the systematics, palaeoecology, morphology and natural history of Mooser’s fossils from Aguascalientes, as much of the previously published information requires further re-examination.
1 MATERIALS AND METHODS
1.1 Study area and geological background
The fossil specimens cited here proceed from the fossiliferous localities that extend through the main water streams in the surroundings of Aguascalientes City. Traditionally, El Cedazo Local Fauna includes fossil taxa mainly from El Cedazo, San Francisco, Cobos and Pargas creeks that comprise a network of water streams with Pleistocene occurrences (Fig. 2).
The geology of the central region of Aguascalientes (which includes the fossil localities) is described by Hernández-Láscares (1979); sedimentation extends from layers with an age prior to the Miocene (possibly Oligocene) to the Holocene (Ferrusquía-Villafranca, 2003; Hernández-Láscares, 1979; Montellano-Ballesteros, 1990). The Ojo Caliente Rhyolite (pre-Miocene) is the oldest lithostratigraphic unit in the region, composed of reddish to brownish-pinkish rhyolites (Ferrusquía-Villafranca, 2003; Hernández-Láscares, 1979). El Zoyatal Tuff (Miocene age) is discordantly overlaid above the Ojo Caliente Rhyolite; it is composed of pyroclastic rocks with brown yellowish to dark grey colouration, and interbedded with fine to medium-grained, arkosic sandstones of fluvial origin, from which the El Zoyatal fossil material was collected (Ferrusquía-Villafranca, 2003; Hernández-Láscares, 1979). El Zoyatal Tuff was deposited because of the strong geological activity that modified the physiography of the national territory during the Miocene (Jiménez-Hidalgo et al., 2002). The Aguascalientes Tuff dates from the Quaternary period and is discordantly arranged above the Miocene sediments; outcrops of this layer are exposed by the erosion in the El Cedazo, San Francisco, Pargas and Cobos creeks, as well as in some water streams near the main localities (Hernández-Láscares, 1979).
Montellano-Ballesteros (1990) described the composition of the Pleistocene localities in Aguascalientes. The fossiliferous sites comprise a sequence of sedimentary deposits with variable lithology and fossil material. The lowermost unit is a lithified grey to beige sandstone with clay and silt-filled fractures, exposed by erosion at Arroyo El Cedazo and Arroyo Cobos. No fossil material was reported in this stratum, and the age is unknown (Montellano-Ballesteros, 1990).
Overlying it is a reddish-brown tuffaceous sand with lenticular conglomerates composed of sub-rounded rhyolitic clasts. This thick unit (up to 30 m) has yielded most of the fossil material, though the preservation is often incomplete. In some areas, angular clasts in a sandy matrix also contain fragmentary remains, and sedimentary structures such as cross-stratification are present; based on biostratigraphy, an Irvingtonian age is attributed to this stratum (Montellano-Ballesteros, 1990).
A discontinuous volcanic ash layer (up to two m thick) overlies parts of this unit and is visible at Arroyo Pargas and some adjacent water streams. Above, a fine whitish sandstone with subangular gravel lenses occurs at Arroyo El Cedazo. Biostratigraphic evidence suggests a Rancholabrean age for this stratum (see Montellano-Ballesteros, 1990), and the age is supported by the fossil occurrences of bison (Mooser & Dalquest, 1975b), as it is a main indicator taxon of this age (Savage, 1951). A dark brown sandy silt lies above, widespread across the region (at Arroyo El Cedazo and Arroyo Pargas, it includes a thin volcanic ash bed); this unit has been exploited for brick production, and some fossil material was recovered (for schemes and detailed information on the stratigraphy of the localities, see Montellano-Ballesteros, 1990; for photos of the fossil sites, see Guzmán-Gutiérrez & Rodríguez-Ávalos, 2008).
2 Examined material
Bibliographic sources that cite some of the material collected by Mooser were consulted to infer the scientific collections in which the fossil specimens from Aguascalientes were deposited (Díaz-Sibaja, 2018; Montellano-Ballesteros, 1990; Prothero et al., 2023; Reynoso-Rosales & Montellano-Ballesteros, 1994; Rincon et al., 2012; see Stevens, 1977; Tapia-García & Sandoval-Ortega, 2024a, 2024b; Vlachos, 2018). Some literature and other documents with information about Mooser were retrieved from the Archivo Histórico del Estado de Aguascalientes (see Estrada, 1989).
Institutional abbreviations. The examined fossil material is deposited in the following scientific collections: Colección Zoológica de la Universidad Autónoma de Aguascalientes (CZUAA, the cited material follows the catalogue number registered by the Instituto Nacional de Antropología e Historia; INAH); Museo Regional de Historia de Aguascalientes (MRHA); Colección Nacional de Paleontología, Museo María del Carmen Perrilliat M., Instituto de Geología, Universidad Nacional Autónoma de México (UNAM), under the acronym “IGM”; and the Texas Vertebrate Paleontology Collections, Jackson School Museum of Earth History, The University of Texas at Austin, under the acronym “TMM”.
Anatomical assessment. Fossil specimens that required a redescription were examined to complement and amend the previous description provided by the authors. Furthermore, in the case of other specimens that were neither characterised nor figured, the fossil material was subjected to a general review conducted through comparison with other specimens reported in literature (Hodnett et al., 2009; Reynolds et al., 2023; Ruiz-Ramoni, 2016).
Abbreviations for dental material. I, upper incisive; P, upper premolar; M, upper molar; c, lower canine;** p,** lower premolar; m, lower molar. The number that follows the dental abbreviation means the position of the tooth.
Stratigraphy and biochronology. International Commission on Stratigraphy (ICS) (2025) periods and epochs are followed, as well as the North American Land Mammal Ages (NALMA) proposed by Savage (1951). The abbreviations for biochronology are as follows: My, million years; ky, thousand years; BP, before Present.
3 RESULTS
A total of 76 fossils collected by Mooser were traced (Tab. 1). Fourteen original fossils of Mooser are deposited in the Colección Zoológica de la Universidad Autónoma de Aguascalientes, four of them newly reported in this work, and 17 in the Colección Nacional de Paleontología, Museo María del Carmen Perrilliat M., Instituto de Geología, UNAM. As for the Vertebrate Paleontology Collections, Jackson School Museum of Earth History, The University of Texas at Austin, more than 500 fossils from Aguascalientes were found. However, much of this collection still requires curation to be properly catalogued, so only 45 specimens were listed, 16 of which are newly reported. Nine holotypes of mammal species described by Mooser and Dalquest were located; some of these, along with additional previously undescribed or unfigured specimens, are addressed in the following section.
TABLE 1 – Traced fossils collected by Oswaldo Mooser. The locality and date are known from notes marked by Mooser on their surface. Names that are currently considered junior synonyms are shown in quotation marks.
| Locality | Family | Species | Referred Material | Reference |
|---|---|---|---|---|
| ORDER ARTIODACTYLA | ||||
| Ce | Antilocapridae | Capromeryx cf. mexicana Furlong, 1925 | IGM 5267, jaw with dental material | Cited as IGM 56-194 by Mooser (1958) |
| Ce | Antilocapridae | Stockoceros conklingi (Stock, 1930) | TMM 42428-9926, skull with horn core base | Mooser and Dalquest (1975b) |
| Ce | Antilocapridae | Stockoceros conklingi (Stock, 1930) | IGM 5268, headgear and cranium | Mooser (1958); Mooser and Dalquest (1975b) |
| SF | Antilocapridae | Tetrameryx sp. | TMM 42428-9882, left jaw fragment | New report, collected in 1964 |
| SF | Antilocapridae | Tetrameryx sp. | TMM 42428-9883, right jaw fragment | New report, collected in 1964 |
| SF | Antilocapridae | Tetrameryx sp. | TMM 42428-9884, right jaw fragment | New report, collected in 1964 |
| SF | Antilocapridae | Tetrameryx sp. | TMM 42428-9885, left jaw fragment | New report, collected in 1964 |
| Pa | Antilocapridae | Tetrameryx sp. | TMM 42428-9887, jaw fragment | New report, collected in 1964 |
| *FC | Antilocapridae | Tetrameryx sp. | TMM 42428-9888, isolated lower tooth | New report |
| SF | Antilocapridae | Tetrameryx mooseri Dalquest, 1974 | TMM 42428-9921, holotype, posterior cranium | Dalquest (1974); Mooser and Dalquest (1975b) |
| SF | Antilocapridae | Tetrameryx mooseri Dalquest, 1974 | TMM 42428-3, isolated dental material | Dalquest (1974); Mooser and Dalquest (1975b) |
| SF | Antilocapridae | Tetrameryx tacubayensis Mooser and Dalquest, 1975 | TMM 42428-2, holotype, cranium with headgear | Mooser and Dalquest (1975b) |
| SF | Antilocapridae | Tetrameryx tacubayensis Mooser and Dalquest, 1975 | TMM 42428-9924, skull | Mooser and Dalquest (1975b) |
| SF | Bovidae | Bison sp. | 1PMP00017496, thoracic vertebra | Tapia-García and Sandoval-Ortega (2024), collected by Mooser in 1977 |
| SF | Bovidae | Bison sp. | 1PMP00017497, thoracic vertebra | Tapia-García and Sandoval-Ortega (2024), collected in 1978 |
| SF | Bovidae | Bison aguascalentensis Mooser and Dalquest, 1975 | TMM 42428-7, holotype, partial cranium | Mooser and Dalquest (1975b) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-26, holotype, mandible | Dalquest and Mooser (1974); Stevens (1977); Prothero et al. (2023) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-19, isolated right m3 | Dalquest and Mooser (1974); Stevens (1977) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-25, fragmentary skull remains | Dalquest and Mooser (1974) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-30, isolated right m2 | Stevens (1977) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-18, dentary with tooth m1, m2 | Dalquest and Mooser (1974) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-21, fragmentary left maxilla | Dalquest and Mooser (1974) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-15, unidentified limb bone | Dalquest and Mooser (1974) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-16, hindlimb remains | Dalquest and Mooser (1974); Stevens (1977) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-14, right dentary, fragmentary | Stevens (1977) |
| Zo | Camelidae | Aguascalientia wilsoni (Dalquest and Mooser, 1974) | TMM 41536-3, two astragalus bones | Dalquest and Mooser (1974) |
| *FC | Camelidae | Camelops sp. | TMM 42428-9823, right maxilla fragment | Mooser and Dalquest (1975b) |
| *FC | Camelidae | Camelops sp. | TMM 42428-9826, right jaw fragment | New report in this work |
| SF | Camelidae | Camelops sp. | TMM 42428-9829, jaw fragment | New report, collected in 1965 |
| Ce | Camelidae | Camelops sp. | TMM 42428-9830, fragmentary left jaw | New report, collected in 1965 |
| SF | Camelidae | Camelops sp. | TMM 42428-9831, fragmentary left maxilla | New report, collected in 1965 |
| SF | Camelidae | Camelops sp. | TMM 42428-9832, left maxilla fragment | New report, collected in 1960 |
| Ce | Camelidae | Camelops sp. | TMM 42428-9834, upper left toothrow | New report in this work |
| SF | Camelidae | Camelops cf. hesternus (Leidy, 1873) | TMM 42428-9824, left jaw fragment | New report in this work |
| Ce | Camelidae | Camelops hesternus (Leidy, 1873) | TMM 42428-9821, mandible | Mooser and Dalquest (1975a, 1975b) |
| Ce | Camelidae | Camelops hesternus (Leidy, 1873) | TMM 42428-9825, right jaw fragment | New report, collected in 1956 |
| Ce | Camelidae | Camelops hesternus (Leidy, 1873) | TMM 42428-9827, jaw fragment | New report in this work |
| ORDER CARNIVORA | ||||
| SF | ND | ND | 1PMP00017484, sacral/coxal vertebra | New report, collected in 1956 |
| SF | Canidae | Aenocyon dirus (Leidy, 1858) | TMM 42428-9781, left dentary | Mooser and Dalquest (1975b) |
| *FC | Canidae | Aenocyon dirus (Leidy, 1858) | TMM 42428-9782, left dentary | Mooser and Dalquest (1975b) |
| SF | Canidae | Aenocyon dirus (Leidy, 1858) | TMM 42428-9783, right dentary | Mooser and Dalquest (1975b) |
| *FC | Canidae | Aenocyon dirus (Leidy, 1858) | TMM 42428-9784, left dentary | Mooser and Dalquest (1975b) |
| *FC | Canidae | Aenocyon dirus (Leidy, 1858) | TMM 42428-9788, right tibia | Mooser and Dalquest (1975b) |
| Ce | Canidae | Canis cedazoensis Mooser and Dalquest, 1975 | TMM 42428-8, holotype, fragmentary right maxilla | Mooser and Dalquest (1975b); Tedford et al. (2009) |
| Ce | Canidae | Canis cedazoensis Mooser and Dalquest, 1975 | TMM 42428-4, fragmentary hindlimb remains | New report in this work |
| SF | Felidae | Panthera atrox (Leidy, 1853) | TMM 42428-9793, radius bone of the right forelimb | Mooser and Dalquest (1975b) |
| Ce | Felidae | Panthera onca (Linnaeus, 1758) | IGM 5253, fragmentary cranial remains | Mooser (1958); Mooser and Dalquest (1975b); Ruiz-Ramoni et al. (2020) |
| ORDER PERISSODACTYLA | ||||
| Ce | Equidae | Equus sp. | 1PMP00017485, fragmentary acetabulum/pelvis | Tapia-García and Sandoval-Ortega (2024), collected in 1960 |
| Ce | Equidae | Equus sp. | 1PMP00017486, fragmentary acetabulum/pelvis | Tapia-García and Sandoval-Ortega (2024), collected in 1960 |
| SF | Equidae | Equus sp. | 1PMP00017571, fragmentary acetabulum/pelvis | Tapia-García and Sandoval-Ortega (2024) |
| Ce | Equidae | Equus sp. | 1PMP00017577, auditory meatus and tympanic bulla | New report, collected in 1975 |
| SF | Equidae | Equus sp. | 1PMP00017487, fragmentary acetabulum/pelvis | Tapia-García and Sandoval-Ortega (2024), collected in 1967 |
| SF | Equidae | Equus sp. | 1PMP00017488, fragmentary acetabulum/pelvis | Tapia-García and Sandoval-Ortega (2024), collected in 1967 |
| Ce | Equidae | Equus excelsus (Leidy, 1858) | TMM 42428-13, cranial remains, holotype of “Onager hibbardi” | Mooser (1958); Mooser and Dalquest (1975b); Reynoso-Rosales and Montellano-Ballesteros (1994) |
| SF | Equidae | Equus cf. conversidens (Owen, 1869) | 1PMP00017536, isolated lower dental material | New report in this work |
| SF | Equidae | Equus cf. conversidens (Owen, 1869) | 1PMP00017543, isolated lower dental material | New report in this work |
| SF | Equidae | Equus cf. conversidens (Owen, 1869) | 1PMP00017498, fragmentary acetabulum/pelvis | Tapia-García and Sandoval-Ortega (2024), collected in 1978 |
| SF | Equidae | Equus cf. conversidens (Owen, 1869) | 1PMP00017526, seventh cervical vertebra | Tapia-García and Sandoval-Ortega (2024), collected in 1981 |
| *FC | Equidae | Equus conversidens (Owen, 1869) | TMM 42428-30, cranial remains with dental material | Mooser and Dalquest (1975b) |
| SF | Equidae | Equus conversidens (Owen, 1869) | IGM 5254, isolated upper teeth | Mooser (1958); Reynoso-Rosales and Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5255, fragmentary jaw remains | Mooser (1958); Reynoso-Rosales and Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5256, isolated upper teeth | Mooser (1958); Reynoso-Rosales and Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5257, right jaw, holotype of “Onager zoyatalis” | Mooser (1958); Reynoso-Rosales and Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5258, fragmentary palatal region | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5259, fragmentary jaw remains | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5260, fragmentary jaw | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5261, fragmentary jaw | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| SF | Equidae | Equus conversidens (Owen, 1869) | IGM 5262, isolated tooth, holotype of “Onager oviedoi” | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens (Owen, 1869) | IGM 5263, fragmentary maxilla | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| SF | Equidae | Equus conversidens (Owen, 1869) | IGM 5264, fragmentary jaw | Mooser (1958); Reynoso-Rosales & Montellano-Ballesteros (1994) |
| Ce | Equidae | Equus conversidens | IGM 5265, isolated lower teeth | Mooser (1958); Reynoso-Rosales (1994) |
| Ce | Equidae | Equus conversidens | IGM 5266, isolated upper teeth | Mooser (1958); Reynoso-Rosales (1994) |
| ORDER PILOSA | ||||
| SF | cf. Megatheriidae | ND | 1PMP00017490, lumbar vertebra | Tapia-García & Sandoval-Ortega (2024) |
| Ce | Mylodontidae | Paramylodon harlani | IGM 5252, hindlimb bone | Mooser (1958); Carbot-Chanona (2021) |
| Ce | Nothrotheriidae | Nothrotheriops cf. shastensis | TMM 42428-9770, cheek tooth | Mooser & Dalquest (1975b) |
| SF | Nothrotheriidae | Nothrotheriops cf. shastensis | TMM 42428-9771, ungual phalanx | Mooser & Dalquest (1975b) |
Abbreviations: Ce: Arroyo El Cedazo (Pleistocene); SF: Arroyo San Francisco (Pleistocene); Pa: Arroyo Pargas (Pleistocene); Zo: El Zoyatal (Miocene); *FC: El Cedazo Local Fauna (Pleistocene), locality not specified; ND: Not determined species.
4 SYSTEMATIC PALEONTOLOGY
Order CARNIVORA Bowdich, 1821
Suborder CANIFORMIA Kretzoi, 1943
Family CANIDAE Fischer von Waldheim, 1817
Subfamily CANINAE Fischer von Waldheim, 1817
Genus Canis Linnaeus, 1758
Type species. Canis familiaris Linnaeus, 1758, by original designation. Middle Pleistocene (around 400 ky BP; for wild ancestors) to Present (Gaubert et al., 2012; Iurino et al., 2022). Formerly Holarctic and Indian (wild ancestors), currently found worldwide due to domestication.
Canis cedazoensis Mooser and Dalquest, 1975b
Figures 3.1–4
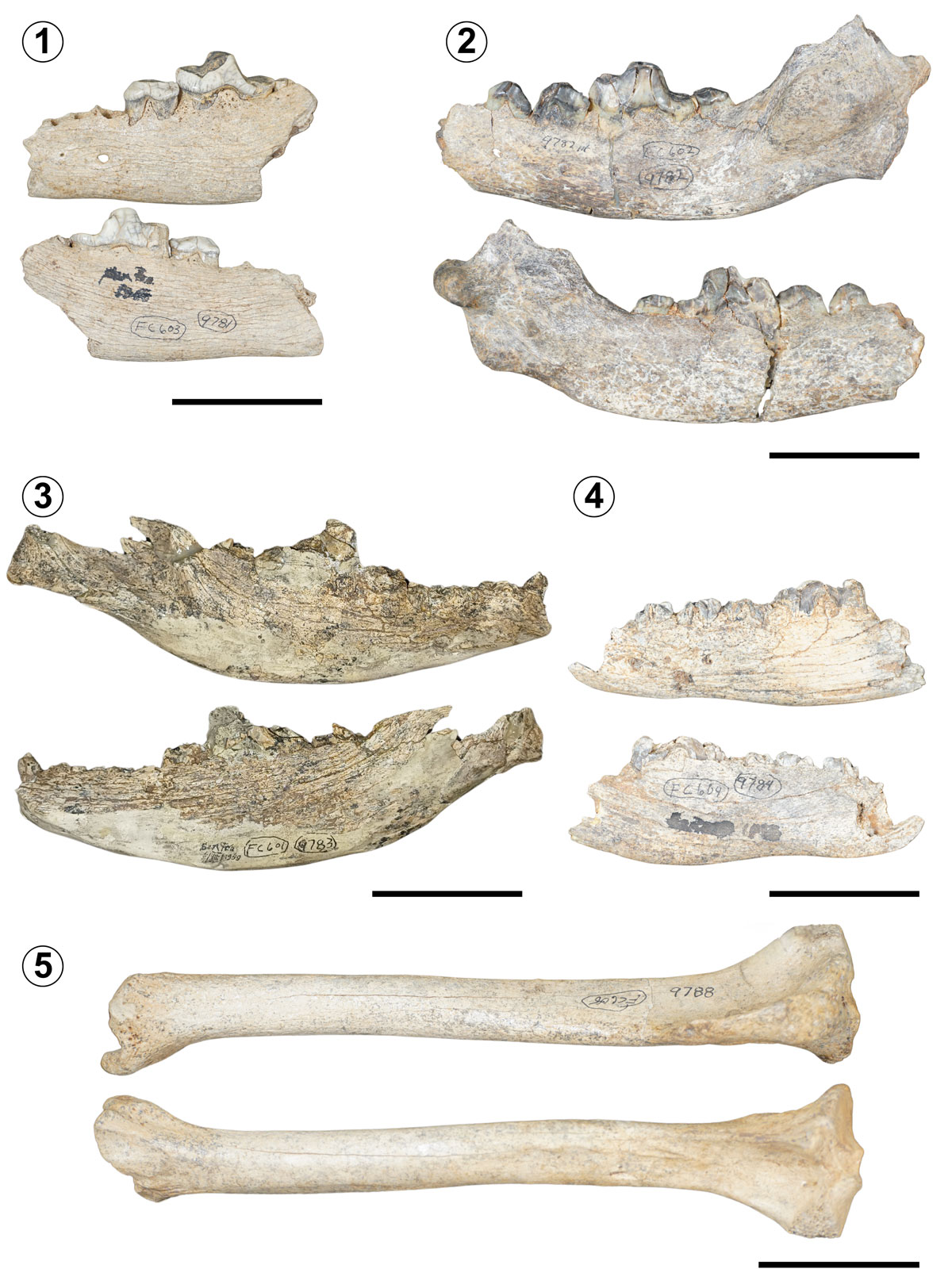
Type Material. TMM 42428-8 (holotype), fragmentary right maxilla with P3, P4 and M1 teeth, with M2 alveolus (Fig. 3.1-3).
Referred Material. TMM 42428-4, metacarpal III, IV and V, with fragmentary remains of phalanges (Fig. 3.4).
Geographic distribution. Currently known only from the type locality, Arroyo El Cedazo, three kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico (Arroyo-Cabrales & Carranza-Castañeda, 2009; Mooser & Dalquest, 1975b).
Stratigraphic range. Known only from the Pleistocene, possibly Irvingtonian/Rancholabrean age (Tedford et al., 2009).
Description. The holotype is fragmentary, but the dental structure remains very complete despite the occlusal surface wear: the main cusp of P3 is flattened and very worn; the P4 exhibits some wear above, and the protocone is missing; the M1 also shows a slightly worn surface. The alveolus of M2 suggests a noticeably smaller tooth size compared to M1. The maxilla presents an infraorbital foramen right above P3 (for morphometric measurements, see Mooser & Dalquest, 1975b). The holotype has a beige-like colouration. The referred material (TMM 42428-4) is in good preservation overall: the metacarpals are structurally complete, and the phalanges are partially complete with some fragmentary elements. The colouration of the referred specimen is noticeably darker compared to the holotype.
Remarks. The authors did not describe additional information on the stratigraphic position of the fossil remains of the species within the locality, which makes it difficult to assign a precise chronological range (Mooser & Dalquest, 1975b). Therefore, the chronostratigraphic information of this taxon remains limited until the discovery of new specimens. According to Mooser and Dalquest (1975b), aside from the holotype, a previously collected and more complete specimen was sent to the United States of America for identification. However, this material never returned, and its current location remains unknown. The referred specimen (TMM 42428-4) represents the first report of postcranial material for this species, as no previous publications have cited or mentioned it.
Discussion. Mooser and Dalquest (1975b) described C. cedazoensis as a small dog, with bigger morphological proportions than the largest North American foxes but smaller than the coyote Canis latrans Say, 1823. Apart from the size, dental morphology is used to diagnose C. cedazoensis: a relatively small M1 compared to the coyote; shorter anteroposterior length of M1 relative to the length of P4, with reduced hypocone and labial cingula in comparison to Canis aureus Linnaeus, 1758 and Canis thooides Tedford, Wang and Taylor, 2009, and a smaller tooth size for M2 compared to M1, inferred by the alveolus size of M2 (Mooser & Dalquest, 1975b; Tedford et al., 2009).
All the teeth measurements (anteroposterior or mesiodistal crown length and buccal-lingual crown width) reported by Mooser and Dalquest (1975b) are smaller in comparison with the ones reported for 18 recent coyotes from Texas by Dalquest (1969). However, compared to the abnormal specimen UMMP 45222 conferred to C. latrans by Fine (1964), C. cedazoensis has a smaller P3 but a more robust P4 in both length and width. In comparison with the red fox Vulpes vulpes (Linnaeus, 1758) measurements from Gingerich and Winkler (1979), C. cedazoensis has bigger morphological dentition in P3 and P4 in both length and width, as well as an M1 with greater length but slightly smaller width. The dentition of C. cedazoensis differs greatly from other fox lineages, such as Lycalopex gymnocercus (Fischer, 1814) (for dental measurements, see Prevosti & Lamas, 2006). Although C. cedazoensis seems to show a distinctive dentition, the holotype requires to be subjected to an extensive systematic and morphometric study to determine if this is a valid species, as the differences in dental morphology may not be sufficiently distinguishable from other canid relatives (for the use of craniodental traits in phylogenetic hypotheses of canids, see Prevosti, 2010).
In their extensive phylogenetic analysis of North American fossil canids, Tedford et al. (2009) mentioned that the known material of C. cedazoensis is too scarce to infer its evolutionary relationships, but due to its morphological features that largely resemble those of the jackal-like canid lineage, it is suggested that it belonged to the same phyletic line. Nevertheless, the systematics of this North American fossil canid lineage is far from being resolved (Prassack & Walkup, 2022), and since the information about this species remains very limited, any known vestige is extremely relevant.
Based on the holotype information provided by Mooser and Dalquest (1975b), some ecological speculations were made, such as the fact that C. cedazoensis possibly occupied a similar ecological niche to that of Old World jackals (Tedford et al., 2009). The relatively large size and particular shape of its dentition may indicate a hypercarnivorous diet, defined as more than 70% composed of meat derived from other vertebrates (Balisi & Van Valkenburgh, 2020), through both predation and scavenging (Bravo-Cuevas et al., 2017). Still, no dental wear analyses have been made to define its dietary ecology, and no coprolites referable to the species have been found to date, in contrast to other Pleistocene canids in Mexico (Bravo-Cuevas et al., 2017), so the essentially carnivorous diet considered by Tedford et al. (2009) and Bravo-Cuevas et al. (2017) also requires a dedicated study.
C. cedazoensis is one of the least studied carnivore species of the Pleistocene in Mexico, which could be attributed to the lack of information available regarding its current location prior to this study. So far, only Tedford et al. (2009) have cited the holotype of C. cedazoensis as TMM 41536-41. However, after the consultation of the material deposited in the Texas Vertebrate Paleontology Collections, The University of Texas at Austin, it was confirmed that the collection series catalogued as “41536” is attributed to Miocene fossils from Aguascalientes, while the Pleistocene specimens are catalogued under the series number 42428, and the actual catalogue number of C. cedazoensis holotype is TMM 42428-8.
Genus Aenocyon Merriam, 1918
Type species. Canis dirus Leidy, 1858, by original designation. Middle to Late Pleistocene (ca. 250 to 12.8 ky BP) (Gedman et al., 2025; Hill et al., 2025; Ruiz-Ramoni et al., 2022). Widespread across North and South America, from Alberta (Canada) to the Argentine Pampas (Prevosti, 2023; Ruiz-Ramoni & Montellano-Ballesteros, 2019).
Aenocyon dirus (Leidy, 1858)
Figures 4.1–5
For a complete synonym list, see Tedford et al. (2009), Ruiz-Ramoni et al. (2022) and Prevosti (2023).
Referred Material. TMM 42428-9781, left dentary with alveolus of p2 and p3, with p4, m1 and m2 teeth (Fig. 4.1); TMM 42428-9782, left dentary with alveolus of p2, with p3, p4, m1 and m2 teeth (Fig. 4.2); TMM 42428-9783, right dentary with p1, p2, p3, p4, m1 and m2 (Fig. 4.3); TMM 42428-9784, left dentary with p2, p3, p4 and m1 (Fig. 4.4); TMM 42428-9788, a well-preserved right tibia (Fig. 4.5).
Geographic distribution. The northernmost records come from two localities in Alberta (Canada); its distribution extends throughout a substantial number of localities across the United States of America, Mexico (Ruiz-Ramoni & Montellano-Ballesteros, 2019), and additional occurrences in South America: Venezuela, Ecuador, Peru (Caro et al., 2022; Ruiz-Ramoni & Montellano-Ballesteros, 2019), Bolivia (Ruiz-Ramoni et al., 2022), possibly northern Chile [based on a conferred specimen; Caro et al. (2022)], and the southernmost record, from the Argentine Pampas (Prevosti, 2023). A recently reported specimen from northeastern China is the first record of A. dirus in Eurasia (Lu et al., 2021). However, some authors do not consider that the morphology and size of the specimen are conclusive evidence for the presence of this species in the Old World (see Ruiz-Ramoni et al., 2022). Specimens collected by Mooser are from Arroyo San Francisco, four kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico (Mooser & Dalquest, 1975b). The precise locality was not specified for each fossil, but in TMM 42428-9781 and TMM 42428-9783, Mooser handwrote the locality on the surface of the specimens.
Stratigraphic range. Middle Pleistocene (ca. 250 ky BP) to Late Pleistocene (ca. 12.8 ky BP) (Gedman et al., 2025; Hill et al., 2025; Ruiz-Ramoni et al., 2022).
Description. Most of the teeth of the specimens are worn, broken, and in some cases even shattered. When Mooser and Dalquest (1975b) mentioned the material referred to A. dirus, the fossil jaws were not properly described. The anatomical evaluation of the teeth and mandibles is presented here for the first time, based on the presence and orientation of the posterior mental foramina, which are situated near or in between p3 and p4 of the external region of the dentary in dire wolves (Hodnett et al., 2009). For the teeth anatomy and position, diagrams of dire wolf mandibles were followed (Reynolds et al. (2023);Figs. 2, 3). The mental foramina are visible in all fossil jaws. In specimen TMM 42428-9781, p4 and m1 teeth are visibly worn on the occlusal surface, while m2 is very fragmentary. In TMM 42428-9782, the dental structure of the premolars and molars is less worn, but noticeably more cracked. Despite the wear and the fissures, these two specimens are the best preserved out of the four dire wolf jaws. The dental material of both specimens, TMM 42428-9783 and TMM 42428-9784, is remarkably broken and shattered. The only measurements and description previously reported were of the right tibia, TMM 42428-9788 (see Mooser & Dalquest, 1975b). This specimen is very complete and preserves the articulation structures, like the medial and lateral intercondylar tubercles and the intercondylar eminence.
Remarks. The dire wolf (A. dirus) is one of the most abundant Pleistocene carnivores in North America (Perri et al., 2021; Ruiz-Ramoni & Montellano-Ballesteros, 2019). This abundance is consistent with El Cedazo Local Fauna records, as the dire wolf is currently the carnivore with the most documented occurrences in Aguascalientes, with at least five different specimens originally collected by Mooser (Fig. 4; Tab. 1). The stratigraphy of these specimens was not documented by Mooser and Dalquest (1975b), so the precise biochronology of this material is uncertain.
Discussion. The species was originally described by Leidy (1854b) under the name Canis primaevus. However, since this name was preoccupied by Hodgson (1833), Leidy (1858) then assigned the name “Canis dirus”. Merriam (1918) suggested that this large canid should be classified into a new genus, Aenocyon, based on morphological evidence, and this recombination is also supported by recent genetic studies (Perri et al., 2021).
A. dirus presents a variety of hypercarnivorous autapomorphies, which are reflected in specific craniodental features. However, it also shares two other synapomorphies with Canis armbrusteri Gidley, 1913 and four with Canis lupus Linnaeus, 1758, but differs from C. lupus by having a more robust skull and teeth (Prevosti, 2023; for Emended Diagnosis, see Tedford et al., 2009). Some diagnostic features of A. dirus are present in the lower dentition (Prevosti, 2023). Nevertheless, as previously noted, most of the teeth in the specimens collected by Mooser are heavily worn or shattered, although some remain viable for more detailed analysis.
Regardless of the abundance of this hypercarnivorous large canid in Aguascalientes, the fossil material of this species collected by Mooser remains poorly studied, since the specimens were never described adequately by the authors. Despite the general description presented here, the morphological and anatomical features of these fossil jaws still need to be complemented by a more dedicated study, which could contribute to the knowledge about this canid lineage in the state territory.
Suborder FELIFORMIA Kretzoi, 1945
Family FELIDAE Fischer von Waldheim, 1817
Subfamily PANTHERINAE Pocock, 1917
Genus Panthera Oken, 1816
Type species. Felis pardus Linnaeus, 1758, by original designation. Early Pleistocene to Present (Paijmans et al., 2018). Northern coasts of Africa and the rest of sub-Saharan Africa, Europe, mainland, south (as far as the Indonesian archipelago) and eastern Asia (Paijmans et al., 2018).
Panthera onca (Linnaeus, 1758)
Figures 5.1, 2
For a complete synonym list, see Seymour (1989) and Manzuetti et al. (2022).
Referred Material. IGM 5253, fragmentary cranial remains of the palatine region and cranial vault: part of the zygomatic arch (jugal bone), right maxilla with P3 tooth and P4 alveolus, along with a part of the premaxilla with I3 (Figs. 5.1, 2).
Geographic distribution. The northernmost fossil records come from the northern United States of America, and they are widely extended throughout most of North, Central and South American regions, to the southernmost region of the Patagonia (Alberdi et al., 2014; Arroyo-Cabrales, 2002; Borrero, 2001; Diaz, 2010). Its current distribution is noticeably limited in comparison to the fossil record and historic distribution, ranging from the southern United States of America, Mexico, Central and South America, to northern Argentina (Diaz, 2010). The specimen collected by Mooser is from Arroyo El Cedazo, at the height of the bridge of the Carretera Federal 70 Aguascalientes–San Luis Potosí, Municipality of Aguascalientes, Aguascalientes, Mexico (Mooser, 1958).
Stratigraphic range. Some fossil remains of P. onca in South America have been found in sites with a possible age of up to one million years BP (Prevosti & Forasiepi, 2018). In North America, the oldest fossil record of the jaguar known to date is from the Hamilton Cave Local Fauna, Pendleton County, West Virginia, initially dated to approximately 850 to 820 ky BP (K. Seymour, 1993), but later considered to be up to 1.6 to 1.3 My (Srigyan et al., 2024). The species survives to the Present (Arroyo-Cabrales, 2002).
Description. The P3 is very worn; its structure is not fragmented, but the cusp shape is not clear because of the tooth wear. The maxilla also presents the alveoli of the protocone and paracone of P4, and the largest alveolus appears to be for the root that is under the metacone of the same P4. Contrary to the description made by Mooser and Dalquest (1975b), the alveolus does not correspond to M1, since felids tend to have a prominent P4 with a greater anteroposterior or mesiodistal length in comparison to M1, but the M1 is considerably wider in buccal-lingual distance than P4 [Ruiz-Ramoni (2016);Fig. 3], so a redescription of the material is presented here. The specimen also has a right infraorbital foramen situated above the space between P3 and P4. The colouration of the specimen is brownish.
Remarks. The specimen IGM 5253 was referred to a jaguar (P. onca) for the first time by Mooser and Dalquest (1975b). However, the authors did not list any anatomic feature that allows to differentiate it from other medium to large-sized Quaternary felids from North America, and the fossil material was not subjected to a proper morphological study, so, although the present study follows the assignation made by Mooser and Dalquest (1975b), it is considered as tentative and in need of a revaluation. Apart from specimen IGM 5253, additional fossil material referred to the Pleistocene jaguar from Aguascalientes was reported by Mooser and Dalquest (1975b), which consists of a tibia and a cervical vertebra (both specimens currently lost).
Discussion. The jaguar is the largest living New World felid (K. L. Seymour, 1989). It resembles the leopard but can be distinguished by its larger size, shorter tail, more muscular build, broader head, and larger, rounder forefoot pads (Nelson & Goldman, 1933). Diagnostic features include distinct craniodental traits and rosette patterns that often contain central black spots (for complete Diagnosis, see K. L. Seymour, 1989).
Mooser (1958) designated the cranial and postcranial material as IGM 56-102 and IGM 56-190, respectively, which implied that, since its discovery, the jaguar of El Cedazo was deposited in the Instituto de Geología, UNAM. Initially, the specimen was identified as Felis sp., since for much of the last century it was common to classify any member of the Felidae within that genus (1912), and Silva-Bárcenas (1969) repeated the taxonomic designation of Mooser (1958) in his list of fossil vertebrates of Mexico.
Years later, Mooser and Dalquest (1975b) mentioned that the big cat fossil from El Cedazo was subjected to a general morphological review, and the authors concluded that it belonged to a jaguar due to the proportions of the maxilla and teeth, which were smaller than those of the big American lion, Panthera atrox (Leidy, 1853), but larger than those of the puma, Puma concolor (Linnaeus, 1771). It is well established that the huge American lion exceeds the jaguar in both dental and body size (Christiansen & Harris, 2009), while pumas exhibit noticeably smaller proportions than jaguars (Torre & Rivero, 2017), despite sharing prey and habitat from Pleistocene to Present (Manzuetti, Jones, Ubilla, et al., 2022).
In their study of a large Pleistocene P. onca specimen found in the state of Jalisco (Mexico), Ruiz-Ramoni et al. (2020) cited the material reported by Mooser. However, the jaguar specimen was not consulted in this study, but its location was inferred based entirely on Mooser’s work (Mooser, 1958; Mooser & Dalquest, 1975b), since even then its location remained unknown. It was not until 2024 that the specimen was rediscovered after decades, when the material from Aguascalientes deposited in the Colección Nacional de Paleontología, Museo María del Carmen Perrilliat M., Instituto de Geología, UNAM, was intensively consulted. However, it was under a new catalogue number (IGM 5253), different from the one initially designed by Mooser (1958), IGM 56-102 (and which is no longer in use), when the El Cedazo jaguar was sent to the Instituto de Geología, UNAM. Additionally, the cranial remains of the jaguar of El Cedazo (now IGM 5253) were still under the name Felis sp., as they had been identified for the first time (Mooser, 1958). The change in catalogue number and the outdated taxonomic designation, along with the limited research on the Aguascalientes fossil fauna, likely contributed to the El Cedazo jaguar being lost for decades.
Mooser (1958) and Mooser and Dalquest (1975b) described the specimen as fragmentary cranial remains of the palatine region, including one incisor and a premolar. This description corresponds with the fossil recovered during the recent consultation (IGM 5253). Additionally, the old catalogue number assigned by Mooser (1958) matches the one written on the surface of the specimen, so it was concluded that it corresponds to the cranial remains of the El Cedazo jaguar. However, as previously noted, the anatomical assessment made by Mooser and Dalquest (1975b) appears to have been inadequate, despite the reanalysis that was mentioned in their study. Furthermore, dental measurements of IGM 5253 were not compared with other jaguar specimens or even other felids.
It is important to note that some Pleistocene jaguar populations in North America developed a body size 15–20% larger than their counterparts that survive today, considered by some authors as an extinct subspecies, Panthera onca augusta (Leidy, 1872) (Anderson, 1984; Carbot-Chanona & Gómez-Pérez, 2014; Ruiz-Ramoni et al., 2020; Simpson, 1941; Slaughter, 1966; Van Devender & al., 2024), and at some point even considered as a different species (Cope, 1880; see Leidy, 1872; McCrady et al., 1951). The size is an important trait, given that body and skull dimensions in extant jaguar populations are often associated with the biomass of available prey (Lavariega & Briones-Salas, 2016).
Specimen IGM 5253 was not properly evaluated with morphometry. While its size suggests that it could belong to a medium to large-sized felid, the identification as P. onca remains to be reassessed. Consequently, an independent and extensive morphometric analysis is necessary to determine whether the specimen belonged to the Pleistocene large-sized jaguar lineage (or even a different phyletic line of Pleistocene felids, since the mere size is not a conclusive taxonomic indicator).
Panthera atrox (Leidy, 1853)
Figures 6.1–4
For a complete synonym list, see Montellano-Ballesteros and Carbot-Chanona (2009).
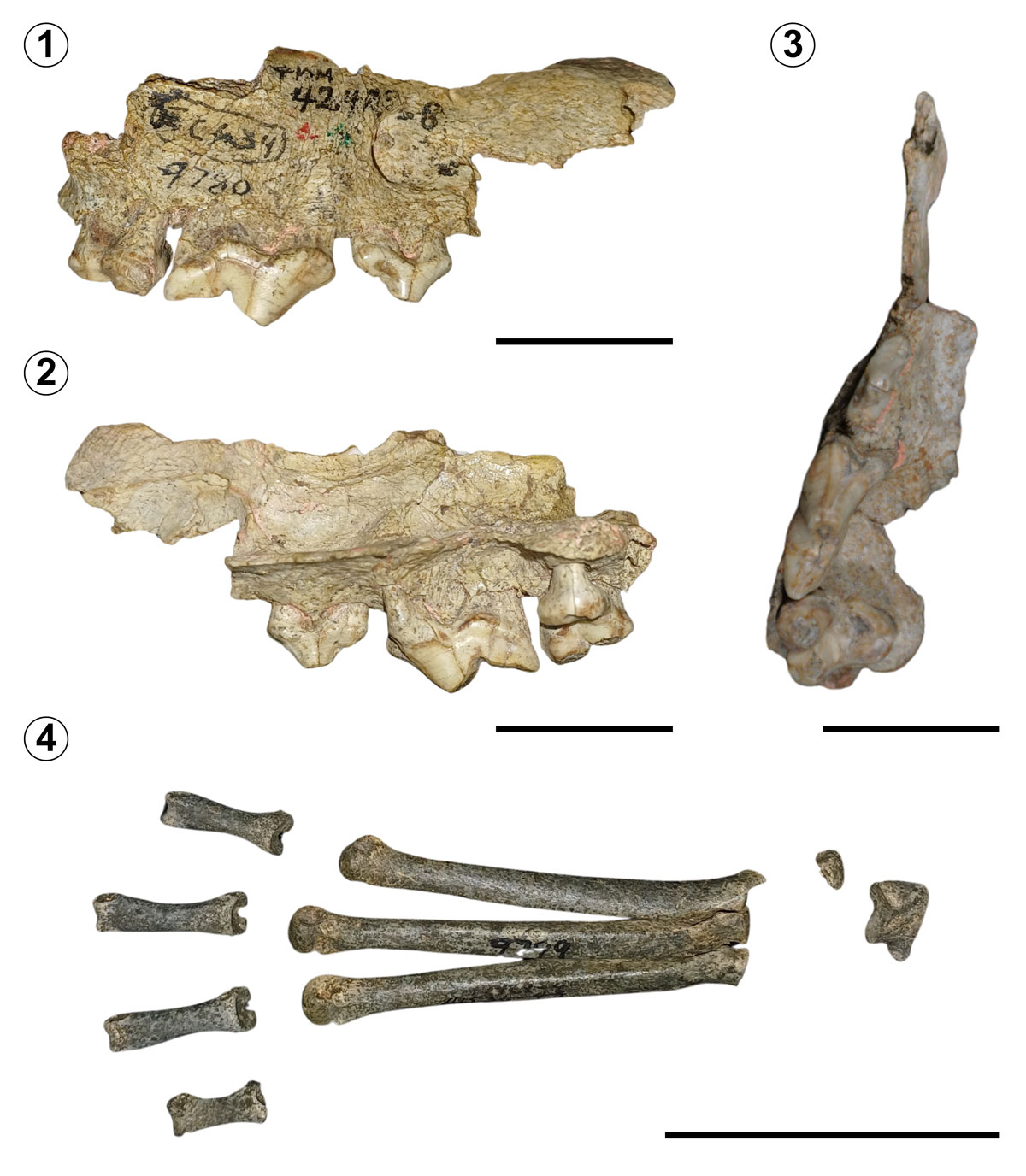
Referred Material. TMM 42428-9793 (Figs. 6,1–4). A large radius bone of the right forelimb.
Geographic distribution. The northernmost record is from Alberta (Canada) (Reynolds et al., 2019), although most of the fossil specimens are from localities found in the United States of America and Mexico (Bravo-Cuevas et al., 2016; Carbot-Chanona & Gómez-Pérez, 2014). The specimen collected by Mooser was found in Arroyo El Cedazo (Pleistocene), three kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico (Mooser & Dalquest, 1975b). The American lion (P. atrox) is broadly accepted as a North American species (Bravo-Cuevas et al., 2016).
However, some authors have considered that a few relatively isolated fossil occurrences of a large felid found in South America could be referred to P. atrox, such as two tentatively referred specimens from the northwest of Peru (Churcher, 1959; Lemon & Churcher, 1961). Additional records from the southernmost regions of the Patagonia have also been interpreted by some authors as P. atrox (Chimento & Agnolin, 2017). However, the fossil material from South America does not provide enough evidence to determine its taxonomic attribution to the American lion.
Stratigraphic range. Ancestral populations of this lineage may have appeared in America during the Middle Pleistocene, giving rise to P. atrox in the Late Pleistocene, since the American lion is considered to be a species mostly limited to the Rancholabrean (Bravo-Cuevas et al., 2016).
Description. The specimen is very well preserved and conserves its main anatomical structures. The head (articular disk) and the neck of the radius, the radial tuberosity, the shaft, the articular surface for carpals and the styloid process remain in very good condition. It has a darker colouration compared to other carnivore fossils from El Cedazo Local Fauna. Measurements of this large specimen were provided by Mooser and Dalquest (1975b).
Remarks. The size of TMM 42428-9793 is unusually large, with a documented total length of approximately 395 mm (for more measurements, see Mooser & Dalquest, 1975b); the taxonomic assessment based on size and anatomical proportions of the radius bone is consistent with those known for the American lion. However, specimen TMM 42428-9793 is slightly larger compared to other reported radii referred to P. atrox (Wheeler & Jefferson, 2009, tab. 4). According to Mooser and Dalquest (1975b), the radius of the American lion from Aguascalientes was only exceeded by a single specimen cited by Merriam and Stock (1932), with a maximum length measured along the internal border of 411 mm.
Discussion. Panthera atrox is a large extinct pantherine, much bigger than any living felid species, distinguished by diagnostic features in its teeth and limb bones (Bravo-Cuevas et al., 2016). The fossil radius of P. atrox found in Aguascalientes has not been studied in detail, possibly due to the poor recovered material that exists, like several Pleistocene felid fossils in Mexico, which tend to be rare and very fragmentary (Bravo-Cuevas et al., 2016; Montellano-Ballesteros & Carbot-Chanona, 2009). As mentioned above, specimen TMM 42428-9793 presents notably larger dimensions compared to other radii subjected to previous analysis; therefore, it is recommended to continue the study of this specimen to evaluate the implications of its large morphotype.
Order ARTIODACTYLA Owen, 1848
Suborder TYLOPODA Illiger, 1811
Family CAMELIDAE Gray, 1821
Subfamily FLORIDATRAGULINAE Maglio, 1966
Genus Aguascalientia Stevens, 1977
Type species. Miotylopus wilsoni Dalquest and Mooser, 1974, by original designation. Known from the type locality, El Zoyatal, Aguascalientes (Mexico), and from the Castolon Local Fauna, Big Bend Texas, USA (Dalquest & Mooser, 1974; Stevens, 1977; Tedford et al., 2004). Early Miocene (Arikareean to Hemingfordian age) (Rincon et al., 2012).
Aguascalientia wilsoni (Dalquest & Mooser, 1974)
Figure 7
Type Material. TMM 41536-26 (holotype), both left and right dentary with teeth (c1 to m3), fragmentary (Fig. 7).
Referred Material. TMM 41536-19, isolated right m3 tooth; TMM 41536-25, fragmentary skull remains with teeth P4, M1 and M2 along a separated jaw with teeth p4, m1 and m2; TMM 41536-30, isolated right m2; TMM 41536-18, dentary fragment with teeth m1 and m2; TMM 41536-21, fragmentary left maxilla with teeth P3, P4; TMM 41536-15, unidentified limb bone; TMM 41536-16, astragalus, partial calcaneus, cuboid, navicular, entocuneiform and proximal ends of metatarsals III, IV and V; TMM 41536-14, right dentary fragment with teeth p2, p3, p4 and m1; TMM 41536-3, two astragalus bones.
Geographic distribution. Currently known only from the type locality, El Zoyatal, four kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico, and a tentatively referred specimen from the Castolon Local Fauna, Big Bend Texas, USA (Dalquest & Mooser, 1974; Stevens, 1977; Tedford et al., 2004).
Stratigraphic range. A Hemingfordian (Early Miocene) age is attributed to the El Zoyatal fossil material (Ferrusquía-Villafranca, 2003), while an Arikareean (Early Miocene) age is estimated for the Castolon Local Fauna specimen (Dalquest & Mooser, 1974; Stevens, 1977; Tedford et al., 2004).
Description. The holotype is fragmentary, and the mandible lies on the sediments in which it was found. Although the left and right dentaries are very fractured, the teeth are structurally complete, slightly worn and of a brachydont type. Additionally, the premolars are relatively short. Dental measurements were reported by Dalquest and Mooser (1974), including a detailed description of the referred material.
Discussion. Aguascalientia wilsoni was described as a small-sized camel with brachydont teeth; similar to Miotylopus gibbi (Loomis, 1911) (=syn. “Miotylopus bathygnathus” Schlaikjer, 1935), but with differences in dental morphology (Dalquest & Mooser, 1974), which also separates it from other Aguascalientia species (Prothero et al., 2023; for Emended Diagnosis, see Rincon et al., 2012). The species was diagnosed within the genus Miotylopus Schlaikjer, 1935, but it was later segregated and included in the genus Aguascalientia Stevens, 1977, of which it is the type species (Stevens, 1977).
Fossils of A. wilsoni are extremely rare among the Miocene fauna of North America, and since Dalquest and Mooser (1974) never mentioned the scientific collection in which the holotype was deposited, the material subsequently remained unaccounted for a long time, until the study of Rincon et al. (2012), which was the first work that specified the scientific collection on which the holotype was housed. Furthermore, Prothero et al. (2023) figured the holotype of A. wilsoni for the first time in decades and clarified that the specimen remained in the Texas Vertebrate Paleontology Collections, The University of Texas at Austin. However, the specimen was cited as TxVP 41536-26, although “TxVP” is one of the many acronyms that were later replaced by the current official acronym for the scientific collection (TMM).
Suborder RUMINANTIA Scopoli, 1777
Family BOVIDAE Gray, 1821
Subfamily BOVINAE Gray, 1821
Genus Bison Hamilton-Smith, 1827
Type species. Bos bison Linnaeus, 1758, by original designation. Holocene, from approximately 5 to 4 ky BP to Present (Carrillo-López et al., 2024; Díaz-Sibaja et al., 2020). From Alberta, Canada, western United States of America to northern Mexico (Farr & White, 2022).
Bison aguascalentensis Mooser & Dalquest, 1975b
Figures 8.1–3
Bison aguascalentensis Mooser and Dalquest, 1975 has been historically considered a synonym of other previously described giant bison species. However, there is no definitive consensus that determines whether it is a synonym of Bison alaskensis Rhoads, 1897 or Bison latifrons (Harlan, 1825). Accordingly, the name assigned by Mooser and Dalquest (1975b) is retained in this study (see discussion for the validity, current status and taxonomic history of the Aguascalientes bison).
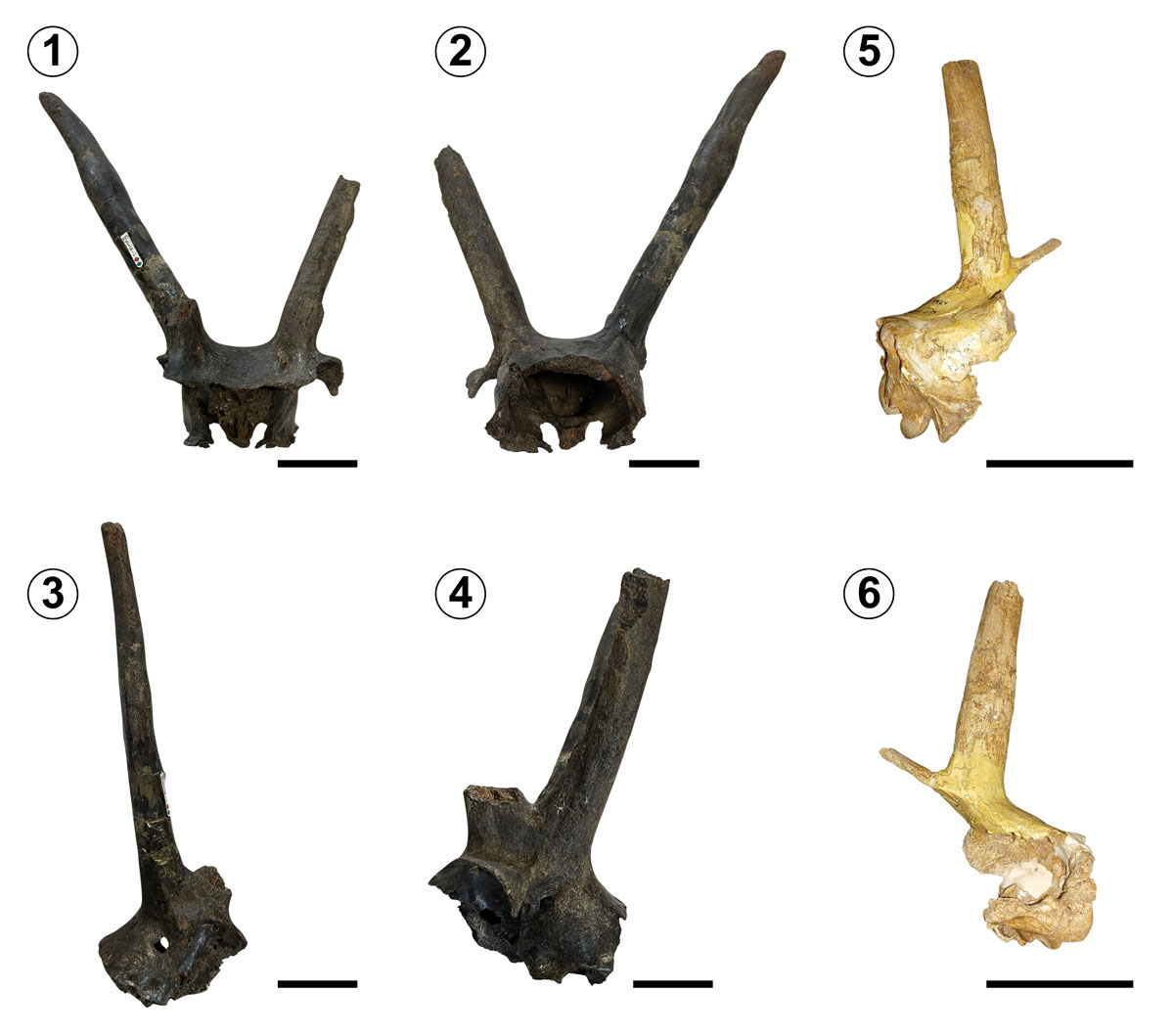
Type Material. TMM 42428-7 (holotype), skull, partial cranium with complete left horn core and basicranium (Figs. 8.1–3).
Geographic distribution. Currently known only from the type locality, Arroyo San Francisco, four kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico (Mooser & Dalquest, 1975b). However, as the taxonomical validity of this species remains uncertain (see discussion below), the distribution of other North American giant bison species is outlined as follows: B. alaskensis, from eastern Beringia, Alaska, Alberta (Canada), most of the United States of America, and central Mexico, where it has been reported in Tajo de Tequixquiac, State of Mexico (Carrillo-López et al., 2024; Pinsof, 1991); B. latifrons, from Alaska, southern Canada, a considerable number of localities across the United States of America, as well as northern, central and southern regions of Mexico, as far as the state of Oaxaca (Carrillo-López et al., 2024; Díaz-Sibaja, 2018; Díaz-Sibaja et al., 2018).
Stratigraphic range. Restricted to the Rancholabrean (Mooser & Dalquest, 1975b; Savage, 1951).
Description. Skull of notably large size, the left horn is almost completely preserved, with only a small portion of the tip missing. The enormous size of the horn may indicate that it is a male individual, since the shape and size of the horn are indicators of sexual dimorphism (Guthrie, 1966), although it is uncertain whether it truly belonged to a male or a female. The anterior region of the skull is absent, while the posterior portion is very complete and relatively well preserved, it conserves the occipital crest and condyles, as well as a very defined foramen magnum.
Remarks. Mooser and Dalquest (1975b) determined that this species corresponds to a lineage of large-sized bison. The horn core was also characterised by being directed at right angle to the longitudinal axis of the skull and noticeably flattened at the base and along much of its length. The authors mentioned that the Aguascalientes bison (B. aguascalentensis) can only be compared to other large bison species, yet, it differs from all of them; the diagnostic feature used by the authors to separate B. aguascalentensis from the most representative species of giant bison, B. latifrons, consists of the very flattened and recurved horn core, in addition to the angle of emergence of the skull close to 90°, which also separates it from B. alaskensis (another large bison species) and related forms, in addition to the shape of the horn core that considerably resembles the shape present in B. alaskensis, but with a much larger size.
Discussion. Bison species are among the largest ruminant mammals in North America (Hernández-Fernández & Vrba, 2005). B. aguascalentensis was a fairly representative taxon of the herbivorous megafauna of El Cedazo (Carreño et al., 1989). However, despite being the type specimen, Mooser and Dalquest (1975b) never specified the scientific collection in which it was deposited, so it was presumed for a long time that the material had been lost (Tapia-García & Sandoval-Ortega, 2024a).
The Aguascalientes bison was described by Mooser and Dalquest (1975b) as a very large-sized bison, characterised by: the growth direction of the horn core, that leaves the longitudinal axis of the skull at a right angle; the distinctively flattened shape of the horn cores at the base and through most of its length; the projection of the horn core after leaving the cranium at a right angle extends slightly downwards, contrary to other bison species in which the horn cores exhibit a pronounced downward curvature (Mooser & Dalquest, 1975b).
The systematics of the Aguascalientes bison has varied over the years: Kurtén and Anderson (1980) recognised B. aguascalentensis as a synonym of B. latifrons, but the authors did not perform a morphological analysis that would demonstrate the synonymy between these two taxa. Later, McDonald (1981) synonymised B. aguascalentensis with B. alaskensis, and Ferrusquía-Villafranca et al. (2010) repeated this taxonomic determination of McDonald for the fossil bison species of Aguascalientes in their list of Mexican Pleistocene fauna. Díaz-Sibaja (2018) discussed part of the description given by Mooser and Dalquest (1975b) for the holotype of the Aguascalientes bison, and argued for the first time the synonymy of B. aguascalentensis with B. latifrons, based on the horn core growth direction and morphology, which allegedly matched the characteristics attributed to B. latifrons, and the reassignment of B. aguascalentensis as a synonym of B. latifrons was proposed. Later, Carrillo-López et al. (2024), based on the study of Díaz-Sibaja (2018), assumed that the bison species that inhabited Aguascalientes during the Late Pleistocene was B. latifrons. This interpretation not only implied a broader distribution area for the giant bison in the Mexican Republic (Carrillo-López et al., 2024; see Díaz-Sibaja et al., 2018) but also contributed to the current understanding that only five Bison species are taxonomically valid for the Quaternary in Mexico (Carrillo-López et al., 2024; Díaz-Sibaja et al., 2020).
Although the argument for the synonymy of B. aguascalentensis with B. latifrons is congruent, the systematic review cannot be conclusive without direct examination of the B. aguascalentensis holotype. Mooser and Dalquest (1975b) provided morphometric information based on the cranial anatomy characters proposed by Skinner and Kaisen (1947), which are used as diagnostic characteristics among Pleistocene bison from North America, but the morphometric indices were erroneously calculated by Mooser and Dalquest (1975b). Now that the location of this specimen is known, a dedicated taxonomic review is required to clarify the systematics of the Pleistocene bison from Aguascalientes.
Some Bison fossils from Aguascalientes are currently deposited at the Colección Zoológica de la Universidad Autónoma de Aguascalientes (Tab. 1). The vertebrae were conferred to Bison antiquus Leidy, 1852, although it is difficult to determine a certain identification with scarce material, so the assignment remains as Bison sp. Another Bison specimen conferred to B. antiquus is deposited at the Museo Regional de Historia de Aguascalientes, which consists of a single M3 tooth (specimen 1PMP00010998 10 358110 1/3). Nevertheless, as mentioned above, taxonomic identification from such little material available for study may not be conclusive. Additional fossil material of Bison sp. in Aguascalientes mentioned by Mooser and Dalquest (1975b) remains to be examined, so the possibility of more than one bison species in Aguascalientes during the Pleistocene is considered.
Family ANTILOCAPRIDAE Gray, 1866
Subfamily ANTILOCAPRINAE Gray, 1866
Genus Tetrameryx Lull, 1921
Type species. Tetrameryx shuleri Lull, 1921. Irvingtonian to Rancholabrean (Davis, 2007; Jarquin-Abundiz et al., 2019). South and southwestern United States of America, northern and central Mexico, down to the state of Puebla (Jarquin-Abundiz et al., 2019).
Tetrameryx mooseri Dalquest, 1974
Figures 9.1–4
Type Material. TMM 42428-9921 (holotype), partial posterior portion of the cranium with both anterior horn cores, the majority of the left posterior and the base of the right posterior horn cores (Figs. 9.1–4) and one M3 associated with the remains of the skull.
Referred Material. TMM 42428-3, isolated dental material (m3).
Geographic distribution. Currently known only from the type locality, Arroyo San Francisco, four kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico (Dalquest, 1974).
Stratigraphic range. Irvingtonian/Rancholabrean age (Davis, 2007; Jarquin-Abundiz et al., 2019).
Description. The holotype preserves much of the headgear structure. The left posterior pronghorn is conserved and presents an oval shape in transversal view; part of both anterior pronghorns is also retained. The presence of both anterior and posterior prongs allows the estimation of the angle of divergence between the tips. The posterior prongs are considerably longer in comparison to the anterior ones, which are short, straight, and thick, with a blunt, flat end. Most of the posterior portion of the cranium is very complete; the occipital crest and condyles are notably well preserved, while the anterior region of the skull is absent.
Remarks. Even though there are many fossils referred to Tetrameryx found in Aguascalientes (Tab. 1), the holotype (TMM 42428-9921) and the referred material (TMM 42428-3) are the only specimens known for T. mooseri that are currently available for study. No postcranial material referred to this species is known to date (Dalquest, 1974; Mooser & Dalquest, 1975b).
Discussion. Dalquest (1974) described T. mooseri as a relatively large four-horned antilocaprid, and the diagnostic features are exclusively found in the headgear: long and slender posterior horn cores; short and stubby anterior horn cores; strongly developed, straight, external sulci confined to the outer surfaces of horn cores; bone of the supraorbital region not expanded into a shelf; horn cores located less prominently over the orbits in comparison to other antilocaprids (Dalquest, 1974). Nevertheless, these traits shall be reconsidered as diagnostic characters, since the headgear morphology might not be sufficiently different from T. shuleri, as Díaz-Sibaja (2018) pointed out.
Tetrameryx tacubayensis Mooser and Dalquest, 1975b
Figure 10.1–6
Type Material. TMM 42428-2, posterior region of the cranium with both bases of anterior horn cores, most of the right posterior horn core, and the base of the left posterior horn core (Figs. 10.1–4).
Referred Material. TMM 42428-9924, skull, dorsocaudal portion with most of the left posterior horn core preserved, fragmentary remains of the left anterior base of the horn core, and a small portion of the posterior base of the horn core (Figs. 10.5, 6).
Geographic distribution. Currently known only from El Cedazo Local Fauna. Type locality: Arroyo San Francisco (Pleistocene), four kilometres southeast of Aguascalientes City. Referred material locality: Arroyo El Cedazo (Pleistocene), three kilometres southeast of Aguascalientes City, Municipality of Aguascalientes, Aguascalientes, Mexico (Mooser & Dalquest, 1975b).
Stratigraphic range. Irvingtonian age (Mooser & Dalquest, 1975b).
Description. The holotype preserves most of both anterior and posterior horn cores, while both anterior appendages conserve only the base of the structure. Although much of the headgear is well preserved, the anterior region of the skull is missing, and the most posterior portion of it is also absent. The type specimen presents a dark colouration, uncommon among the fossils from El Cedazo Local Fauna, in contrast to the specimen TMM 42428-9924, which has a beige colouration with some orange-like regions. In both the holotype and referred material, the angle of divergence between the anterior and posterior horn cores can be estimated. As for the rest of the referred material, the posterior region of the skull is better preserved than in the holotype: the occipital condyles are present, and the foramen magnum is clearly visible.
Remarks. Just as it happens with T. mooseri, the holotype and the referred material of T. tacubayensis are the only fossil specimens assigned to T. tacubayensis that are currently available for study. Apart from the holotype (TMM 42428-2) and the referred material (TMM 42428-9924), Mooser and Dalquest (1975b) mention an additional specimen, which consists of the base of the headgear with some of the anterior and part of a posterior pronghorn, but the specimen is not cited, as the location of the material is unknown. T. tacubayensis is only known from cranial material; there are no appendicular skeleton remains known to date (Mooser & Dalquest, 1975b).
Discussion. The antilocaprid fossils are often characterised by their integumentary appendages, which are described by O’Gara (1990) as unique anatomic structures that are not strictly considered horns or antlers, due to the fact that they have an ossified horn core that is also covered by a sheath that is detached annually, like antlers of cervids. In some antilocaprid lineages, these headgear structures have been historically called “nondeciduous antlers” or “pseudoantlers” (Marriott & Prothero, 2022), but for the lineage of antilocaprines, to which Tetrameryx Lull, 1921, Capromeryx Matthew, 1902 and Stockoceros Frick, 1937 belong (Davis, 2007), the “pronghorn” designation may be the most appropriate due to the evolutionary and anatomical nature of the integumentary appendages (Davis et al., 2011; Marriott & Prothero, 2022). The revised diagnosis of Tetrameryx by Flora (2019) states that Tetrameryx possesses some of the longest prongs among the antilocaprids, along with a pronounced difference in length between the small anterior and the elongated posterior horns.
Mooser and Dalquest (1975b) described T. tacubayensis based on headgear morphology, and the diagnostic traits are as follows: the anterior tines of the horn cores are strongly inclined anteriorly, with a wide divergence angle, similar to Tetrameryx irvingtonensis Stirton, 1939, the anterior horn cores are also shorter than the posterior ones; the pronghorn base is narrow and relatively elevated at the bifurcation (point of divergence of tines); a slender posterior tine, straight and round without external sulcus; the external sulcus is visible on the bases of anterior tines; a well-developed supraorbital shelf; braincase grooved laterally medial to the supraorbital shelf Mooser & Dalquest (1975b).
As discussed before, diagnosis based on headgear morphology in antilocaprids may not be very efficient, since the angle of divergence between prongs varies during ontogenetic development (and even between adult specimens) in this type of antilocaprids (Furlong, 1943); consequently, it is not recommended to use it as a diagnostic feature(Lull, 1921; Marriott & Prothero, 2022). Both T. mooseri and T. tacubayensis were described based on variations in the lateral sulcus of the posterior prong and the angle of divergence between the tips (Figs. 9.3, 10.3,4). Additionally, no other vestige of T. mooseri and T. tacubayensis is known apart from the ones reported in the fossiliferous localities of Aguascalientes to date (Bravo-Cuevas et al., 2013). Given the restricted occurrences of T. mooseri and T. tacubayensis in the fossil localities of Aguascalientes, along with the fact that no Pleistocene localities have been recorded with more than one species of antilocaprid of the same genus, except for El Cedazo Local Fauna (White Jr. et al., 2022), the taxonomic status of both Tetrameryx species is questioned.
The antilocaprid fossils of Aguascalientes remain poorly studied, perhaps due to the fact that the location of the holotypes of the two species has not been documented in literature yet. Bravo-Cuevas et al. (2013) mentioned that when antilocaprid fossils from El Cedazo Local Fauna were reported, some specimens were not adequately described nor were they figured, and more information is needed for further studies. The description of these antilocaprid fossils, as well as the figured material (Figs. 9–11), contributes to the schematisation of these specimens. However, the material remains to be subjected to an extensive systematic and morphometric study, in order to determine the validity of the two Tetrameryx species that lived in Aguascalientes.
Genus Stockoceros Frick, 1937
Type species. Tetrameryx conklingi Stock, 1930, by original designation. Rancholabrean age (Bravo-Cuevas et al., 2013; Davis, 2007). Southern United States of America to eastern and central Mexico (Bravo-Cuevas et al., 2013; Davis, 2007).
Stockoceros conklingi (Stock, 1930)
Figures 11.1–5
For a complete synonym list, see Díaz-Sibaja (2018).
Referred Material. IGM 5268, partial remains of the skull, with most of both anterior and posterior horn cores well preserved (Figs. 11.1–5).
Geographic distribution. Southern United States of America (Arizona and New Mexico), northern, eastern and central states of Mexico (Bravo-Cuevas et al., 2013; Davis, 2007).
Stratigraphic range. Rancholabrean age (Bravo-Cuevas et al., 2013; Davis, 2007).
Description. The specimen preserves most of the headgear structure, including both anterior and posterior horn cores in very good condition; the base of these appendages is complete, and just the tips of the horns are missing. The bifurcated horn cores, which share a common base located above and behind the orbit to some degree, are congruent with a diagnostic feature of the species. The angle of divergence of this specimen is visible since it conserves the cranial base. The cranial vault is very fragmentary; the posterior portion of the skull is broken, and the occipital crest and condyles are missing. This specimen presents a beige-like colouration, like the Rancholabrean fossils from the Pleistocene localities in Aguascalientes, and it belongs to a young adult specimen (Díaz-Sibaja, 2018).
Remarks. Specimen IGM 5268 was initially reported as Tetrameryx sp. with the old catalogue number IGM 56-204 by Mooser (1958). However, Mooser and Dalquest (1975b) reassigned the fossil material to S. conklingi. Although the authors did not specify any diagnostic features that separate IGM 5268 from Tetrameryx, the specimen shows bifurcated horn cores that grow from a shared base, which is a diagnostic character of S. conklingi. Thus, the reassignment of specimen IGM 5268 from Tetrameryx sp. to S. conklingi is followed here based on that diagnostic character.
Out of all the fossil material referred to S. conklingi found in Aguascalientes, specimen IGM 5268 stands out as the most complete cranial material (Mooser & Dalquest, 1975b). Additional skull remains were found, but their location has not been reported yet. Specimen IGM 5268 is the only antilocaprid fossil from Mooser’s collection whose location has been reported; the original catalogue number (IGM 56-204) implied that the specimen was deposited in the Instituto de Geología, UNAM, and Díaz-Sibaja (2018) cited the material with the current catalogue number.
Discussion. Stockoceros was first described by Frick (1937) as a subgenus, and later, Skinner (1942) transferred the name and reassigned it as a different genus. However, even though the author citation of the genus can be found in literature as Skinner, 1942 (Bravo-Cuevas et al., 2013), the original use of the name corresponds to Frick (1937), so in this study it is considered the author citation for Stockoceros.
The type species, S. conklingi, is diagnosed by many headgear characteristics, including bifurcated pronghorns that originate from a shared base, located above and slightly behind the orbit on each side. Additionally, the anterior pronghorns are thicker and longer than the posterior ones, although not as much as in T. shuleri (for complete Diagnosis, see Stock, 1930). These two traits are clearly visible in specimen IGM 5268, so the later reassignment of the material made by Mooser and Dalquest (1975b) is followed.
Mooser (1958) reported the presence of two different taxa of antilocaprids belonging to Capromeryx and Tetrameryx. Later, Mooser and Dalquest (1975b) reported Capromeryx minor Taylor, 1911 [=syn. “Capromeryx mexicana”;Furlong (1925)], both species of Tetrameryx: T. mooseri and T. tacubayensis, and finally, S. conklingi. Nevertheless, the antilocaprid lineage in Aguascalientes is not limited to the extinct Pleistocene species reported by Mooser (1958), Dalquest (1974), Mooser and Dalquest (1975b); archaeozoological remains of the only extant pronghorn species Antilocapra americana (Ord, 1815) have been found in the archaeological site El Ocote (Pérez Ríos, 2023), which is located southwest the Municipality of Aguascalientes, on the eastern slopes of the Sierra Madre Occidental (Pelz Marín, 2023), although this species cannot be found in the state territory in recent times (Chávez-Andrade et al., 2015).
5 DISCUSSION
A considerable number of fossil occurrences in the state of Aguascalientes were documented through the years by Oswaldo Mooser and Walter W. Dalquest. Miocene fossils from Aguascalientes are much less abundant than those dating to the Pleistocene. However, even though El Zoyatal locality has scarce material, the location of these specimens is relatively better documented in comparison to the Pleistocene fauna from the state. The whereabouts of the fossil material from El Zoyatal were never reported by Dalquest and Mooser (1974), but few years later, Stevens (1977) mentioned that Dalquest and Mooser (1974) had catalogued several specimens of El Zoyatal as part of the fossil collection of Midwestern State University, Wichita Falls, Texas (those specimens can be found in literature under the acronyms MSU, MU, or MWSU), and part of the material was ultimately curated in the Texas Vertebrate Paleontology Collections, The University of Texas at Austin (formerly Texas Memorial Museum) (Stevens, 1977), which is the collection where they are currently deposited.
According to collection staff, the Vertebrate Paleontology Collections, The University of Texas at Austin, has incorporated a variety of fossil material that has been cited in published literature under different acronyms such as BEG, TMMNH, TxVP (which was used by Prothero et al., 2023 for the holotype of A. wilsoni), UTBEG, TNSC, UT, VPL, MSU, MU, MWSU, TAM , TAMU, ETSU, ETSC and TAMUK, thus, the reported material with any of those acronyms indicates that it is now part of the Vertebrate Paleontology Collections.
The location of some of the Miocene fossils of Aguascalientes was directly mentioned in literature by Montellano-Ballesteros (1990), although it was through personal communications and no fossil specimens were cited in that study. Reynoso-Rosales and Montellano-Ballesteros (1994) cited some fossils collected by Mooser with the catalogue number of the Texas Vertebrate Paleontology Collections, The University of Texas at Austin, but the referenced material corresponded entirely to Pleistocene horses, and the rest of the material from El Cedazo Local Fauna remained without a specific published location.
It was reported that hundreds of specimens were donated by Mooser to the Instituto de Geología, UNAM (Estrada, 1989). However, the number of fossils collected by Mooser in Aguascalientes that are currently available for study in the Colección Nacional de Paleontología, Museo María del Carmen Perrilliat M. of the Instituto de Geología is too small in comparison to the 275 specimens that were initially donated, and the location of the rest of the material is still unknown.
Some institutions, such as the Museo Regional de Historia de Aguascalientes, have made efforts to recognise the scientific labour made by Mooser. Additionally, the information about the fossil material from Aguascalientes that is housed in this collection has been publicly known for a long time and recently reported in literature (Tapia-García & Sandoval-Ortega, 2024a, 2024b). However, Vlachos (2018) mentioned that the holotype of the tortoise Gopherus auffenbergi Mooser, 1972 found in Arroyo Cedazo had an unknown location, even though the whereabouts of this specimen, along with more turtle and tortoise remains, were reported by Guzmán-Gutiérrez and Rodríguez-Huerta (1993). The fossil material was housed at the Museo Regional de Historia de Aguascalientes (1993), where it remains to this day. The location of a lot of the specimens collected by Mooser in Aguascalientes is unknown to several authors since no work has been done to report the current location of these fossils. The lack of published information about the fossil fauna of Aguascalientes is perhaps the main reason why there were no more records on the current location of this material, as some local authors had even conjectured that several type specimens were still housed at Midwestern State University, Wichita Falls, Texas (Valencia-Cruz & Guzmán-Gutiérrez, 1994).
Even though the majority of the material collected by Mooser comes from Aguascalientes, he collected some specimens in other states of Mexico, such as Guanajuato, from El Ocote locality (some of this material is currently deposited at the Colección Zoológica de la Universidad Autónoma de Aguascalientes). Thus, it is important to continue tracking not only the palaeontological material collected by Mooser that comes from Aguascalientes, but also the rest of the specimens from different localities and states.
This work presents, for the first time, a comprehensive account of the location and general description of a substantial portion of fossils from Aguascalientes. The rediscovery of these specimens holds important implications, not only due to their heritage value (Morales-Ortega et al., 2021) but also because it marks a major step towards establishing a basis for future research. Although much of the fossil material is now compiled, described and discussed here, notable challenges persist, as a considerable portion of Mooser’s collection remains in need of curation, analysis, and documentation. This highlights the necessity for continued palaeontological research and curatorial efforts to ensure that all specimens are completely catalogued and properly evaluated.
5.1 Implications for Pleistocene mammal diversity in Aguascalientes
Among the palaeontological species reported for Aguascalientes, few have been subjected to extensive studies. Previous systematic reviews have been conducted on some Pleistocene mammals of the state, an example of these contributions includes the studies of Pleistocene camels: Mooser and Dalquest (1975a) described Camelops traviswhitei Mooser and Dalquest, 1975 from cranial and mandibular fragments, currently deposited in the Texas Vertebrate Paleontology Collections, The University of Texas at Austin (TMM 42428-9821), along with multiple fossil material referred to the genus. However, a study of morphological variation in Camelops Leidy, 1854 (Leidy, 1854a) showed that C. traviswhitei corresponds to a specimen of western camel Camelops hesternus (Leidy, 1873) with population variation (Baskin & Thomas, 2016), and the species described by Mooser and Dalquest (1975a) is now broadly accepted as a synonym of C. hesternus (Carbot-Chanona et al., 2023; Machorro-Román et al., 2018).
Equids are another example of Pleistocene fauna whose systematics has been extensively reviewed. Horses are arguably the most abundant mammalian lineage from the Pleistocene in Aguascalientes (Mooser, 1958; Mooser & Dalquest, 1975b). Mooser (1958) initially determined the presence of ten species of Pleistocene horses belonging to the genera Onager Brisson, 1762, Asinus Brisson, 1762 and Equus Linnaeus, 1758 for Aguascalientes. Later, Mooser and Dalquest (1975b) reduced the number of horse taxa for the Pleistocene of Aguascalientes to seven, this time grouping all the species within the genus Equus. However, from an ecological approach, it was questionable that many similar horse taxa have coexisted in locality and temporality (Reynoso-Rosales & Montellano-Ballesteros, 1994).
Subsequent morphological and ecological revisions suggested that it is probably just a single species: Equus conversidens (Owen, 1869), since most of the material found from horses in El Cedazo can be referred to this taxon, and the diagnostic traits mentioned by Mooser (1958) and Mooser and Dalquest (1975b) for the other species fall within the range of intraspecific variation of E. conversidens (Montellano-Ballesteros, 1990; Reynoso-Rosales & Montellano-Ballesteros, 1994; Rodríguez-Ávalos, 1991). However, other authors suggested that the dental morphology of fossil horses from El Cedazo evidences the presence of three additional recognised species apart from E. conversidens in Aguascalientes during the Pleistocene: Equus mexicanus Hibbard, 1955, Haringtonhippus francisci (Hay, 1915) (=syn. “Equus francisci” Hay, 1915) and Equus cedralensis Alberdi, Arroyo-Cabrales, Marín-Leyva and Polaco, 2014 (Alberdi et al., 2014; Jiménez-Hidalgo et al., 2024; Jiménez-Hidalgo & Díaz-Sibaja, 2020; Priego-Vargas et al., 2017). The possibility of more than one horse species in the region is plausible, since many specimens deposited in the Texas Vertebrate Paleontology Collections, The University of Texas at Austin, have not been considered in previous studies. Further research on Pleistocene horses of Aguascalientes will clarify this issue.
As mentioned before, some of the new species that Mooser and Dalquest described in Aguascalientes also require more evidence to be considered valid: the taxonomic status of C. cedazoensis has not been questioned yet, but a more robust morphometric study might support its identity; the Aguascalientes bison, B. aguascalentensis, is currently not considered a valid species of bison (Carrillo-López et al., 2024; Díaz-Sibaja, 2018; Díaz-Sibaja et al., 2020; McDonald, 1981), but the forgotten holotype specimen (TMM 42428-7) was never examined in these works, so it should be considered in further studies; at last, the validity of the two species of Tetrameryx (T. mooseri and T. tacubayensis) should also be re-evaluated, as their description might be based on non-diagnosable characters.
Apart from the El Cedazo Local Fauna holotypes, additional fossil material remains to be studied. Few lineages of North American Pleistocene megafauna have survived to the Present (Broughton & Weitzel, 2018), such as the jaguar. As discussed previously, the reassessment of the El Cedazo jaguar (IGM 5253) is necessary, as it corresponds to one of the few specimens referred to P. onca known to date that support the presence of this pantherine lineage in Aguascalientes (Mooser & Dalquest, 1975b). This suggests that the presence of the jaguar in the state was gradually reduced during the Pleistocene until the ancestral populations disappeared. However, a wandering jaguar individual was recently recorded in the Monte Grande locality, Sierra Fría, Municipality of San José de Gracia, in the northwest region of the state (Quintero-Díaz & al., 2024), which will have implications for the study of this species in Aguascalientes. Jaguars are currently threatened by habitat loss due to human activities (Ceballos, J. de la Torre, et al., 2021; Ceballos, Zarza, et al., 2021; Torres-Romero & al., 2023), so the study of this lineage of felids can provide relevant information for understanding its conservation in the region.
Fossil occurrences of carnivores are noticeably less abundant than those of their prey and other mammals, mainly due to their role as predators in food webs (Martin, 1989). However, there are multiple records of carnivorous mammals in Aguascalientes, maybe due to the variety of herbivorous megafauna that lived in the state during the Pleistocene. Mooser and Dalquest (1975b) suggested that the El Cedazo Local Fauna inhabited an open ecosystem of grasslands or prairies, which provided an abundant source of food for the herds of large grazing mammals and therefore, the development of large populations of carnivores. The expansion of the Chihuahuan desert during the Pleistocene was addressed afterwards in a vegetation study (Siqueiros-Delgado et al., 2016), which could also be related to the evidence of extensive grasslands in the past of Aguascalientes.
6 CONCLUSIONS
An extensive systematic review of the fossil specimens from El Cedazo Local Fauna should be carried out in order to corroborate or invalidate the taxonomic status of the unique palaeontological species of the state: Canis cedazoensis requires additional studies that support its current validity, as well as Tetrameryx mooseri and T. tacubayensis, due to limited or ambiguous diagnostic evidence. Bison aguascalentensis holotype shall be considered in upcoming research to determine which bison species lived in Aguascalientes during the Pleistocene. In addition to these unique Pleistocene species, the reassessment of other specimens, such as the El Cedazo jaguar, is required. A broader systematic study of equids is also needed to corroborate the taxonomic status of the reported species, this time incorporating the fossil material of Pleistocene horses available for study in the Vertebrate Paleontology Collections, The University of Texas at Austin. Likewise, now that the location of more fossil specimens from Aguascalientes is known, further palaeoecological studies are intended to be conducted to complement the knowledge of the past biodiversity of Aguascalientes.
At the time, Mooser and Dalquest made meaningful palaeontological contributions in Aguascalientes, but currently, the research on the fossil fauna of the state is very limited. Even though the present work reports a detailed compilation of the location of several specimens that were considered lost, it is still necessary to continue with the exhaustive search for the location of fossils from Oswaldo Mooser’s collection.
7 ACKNOWLEDGEMENTS
I want to thank Jaime Escoto and Mónica Croce Hernández Duque of the Colección Zoológica de la Universidad Autónoma de Aguascalientes (CZUAA) for the permission to consult the material, Laura Verónica Balandrán González, the personnel of the Museo Regional de Historia de Aguascalientes (MRHA), Marco Octavio de Lira Mendoza, Miguel Sánchez-Salinas, José Alberto Cruz-Silva and the Instituto Nacional de Antropología e Historia (INAH) for the curation data and preliminary report of the fossil specimens from local collections. I am grateful to Violeta Amparo Romero-Mayén and Jesús Alvarado-Ortega of the Colección Nacional de Paleontología, Museo María del Carmen Perrilliat M., Instituto de Geología, UNAM, for allowing access to the collection for consultation and review of the fossil material from Aguascalientes. I extend my thanks to James Chris Sagebiel of the Texas Vertebrate Paleontology Collections, Jackson School Museum of Earth History, The University of Texas at Austin, for his invaluable assistance with Oswaldo Mooser’s collection, including access, preparation of the material, curation data, and permission to use the provided photos. Thanks to Manuel Higinio Sandoval-Ortega and Gloria Stephanie Villa-Jaimes for their comments on the preliminary version of the manuscript and their support during the realisation of this project. To Jesús Emilio Ramos-Heredia for his comments on the antilocaprid morphology; José Alberto Rodríguez-Ávalos for giving historical context of Mooser’s contributions, palaeontology in Aguascalientes, and for sharing references on the equids from El Cedazo Local Fauna; Damian Ruiz-Ramoni for his comments and feedback on the anatomical features of the El Cedazo jaguar as well as for sharing his PhD Thesis; José Jesús Sigala-Rodríguez for facilitating the opportunity to hold a scholarship at the Departamento de Biología, Universidad Autónoma de Aguascalientes, which enabled the visits to the scientific collections; Joaquín Chávez Pérez from the Archivo Histórico del Estado de Aguascalientes, for providing important information of Mooser for its use in this work, and Rodrigo A. Medellín, for sharing relevant bibliography for the discussion of the biogeographic history of the jaguar in Mexico. The comments and suggestions of the anonymous reviewers are also greatly appreciated, as is the guidance of the editors María Encarnación Pérez, Maria Eugenia de Porras and Javier González throughout the preparation of this work. Finally, I would like to express my gratitude to Oswaldo Mooser and Walter W. Dalquest for their relevant contributions to the palaeontology knowledge of Aguascalientes.
