
RECORDS OF ATURIA PERUVIANA (NAUTILIDA, CEPHALOPODA) FROM THE SAN JACINTO FORMATION (UPPER EOCENE TO LOWER OLIGOCENE), OVEJAS (SUCRE, COLOMBIA)

New specimens of Aturia (Nautilida) were collected from the late Eocene to early Oligocene deposits of the San Jacinto Formation near Ovejas (Sucre, northern Colombia). These specimens, alongside historical material collected from a nearby region, are identified as Aturia peruviana. The morphological analysis of the shell (the phragmocone and suture line characters) confirms that Aturia colombiana is a junior synonym of A. peruviana, in conformity with previous taxonomic proposals. Furthermore, the stratigraphic range of A. peruviana in Colombia is constrained to the upper Eocene to lower Oligocene, consistent with its documented occurrence in Peru.
Cephalopoda, Eocene–Oligocene, Aturiidae, Aturia, San Jacinto Formation, Colombia, Nautiloidea
Figures


Resumen. REGISTROS DE ATURIA PERUVIANA (NAUTILIDA, CEPHALOPODA) DE LA FORMACIÓN SAN JACINTO (EOCENO SUPERIOR AL OLIGOCENO INFERIOR), OVEJAS (SUCRE, COLOMBIA). Se recolectaron nuevos especímenes de Aturia (Nautilida) en los depósitos del Eoceno tardío al Oligoceno temprano de la Formación San Jacinto, cerca de Ovejas (Sucre, norte de Colombia). Estos especímenes, junto con material histórico recolectado en una región cercana, se identifican como Aturia peruviana. El análisis morfológico de las características de la conchilla (tanto del fragmocono como de la línea de sutura) confirma que Aturia colombiana es un sinónimo menor de A. peruviana, en conformidad con propuestas taxonómicas anteriores. Además, el rango estratigráfico de A. peruviana en Colombia se limita al Eoceno superior–Oligoceno inferior, lo que concuerda con su presencia documentada en Perú.
Palabras clave. - Cephalopoda. - Eoceno–Oligoceno. - Aturiidae. - Aturia. - Formación San Jacinto. - Colombia. - Nautiloidea.
EVEN THOUGH there have been nautilid reports from Colombia, ranging from the Permian to the Miocene, their fossil record is still scarce, with only eight genera identified so far. Domatoceras Hyatt (1891), and Titanoceras? Hyatt (1884), are known from a Permian succession of the Perijá mountain range (Patarroyo et al., 2019; Thompson & Miller, 1949), Oxynautilus? Mojsisovics (1902), and Cenoceras Hyatt (1884), from the upper Triassic of Chaparral (Tolima) (Geyer, 1973; Perilla, 2018), Heminautilus Spath (1927), from the Aptian of San Luis, Tolima (Durham, 1946), Firavitoba (Boyacá) and Villanueva (Santander) (Patarroyo (2001);D. Atuesta pers. comm.), Cymatoceras Hyatt (1884), and Eutrephoceras Hyatt (1894), from the Aptian of Tolima (Durham, 1946), and Aturia Bronn (1838), from the Cenozoic of Bolívar, Sucre and La Guajira (Anderson, 1928; Clark & Durham, 1946; Jung, 1966).
Anderson (1928) reported the first nautilid from Cenozoic deposits of Colombia based on a specimen found in the Sinú—San Jacinto Belt, near El Carmen de Bolívar (Bolívar, Colombia). In 2018, members of the Hydrocarbon Division of the Servicio Geológico Colombiano (SGC), in the framework of the project “Diapiros de Lodos”, collected 13 new nautilid specimens from a nearby region. These specimens allow us to compare and discuss the taxonomy, nomenclatural aspects, and stratigraphic position of these Cenozoic nautilids.
1 GEOGRAPHICAL AND GEOLOGICAL CONTEXT
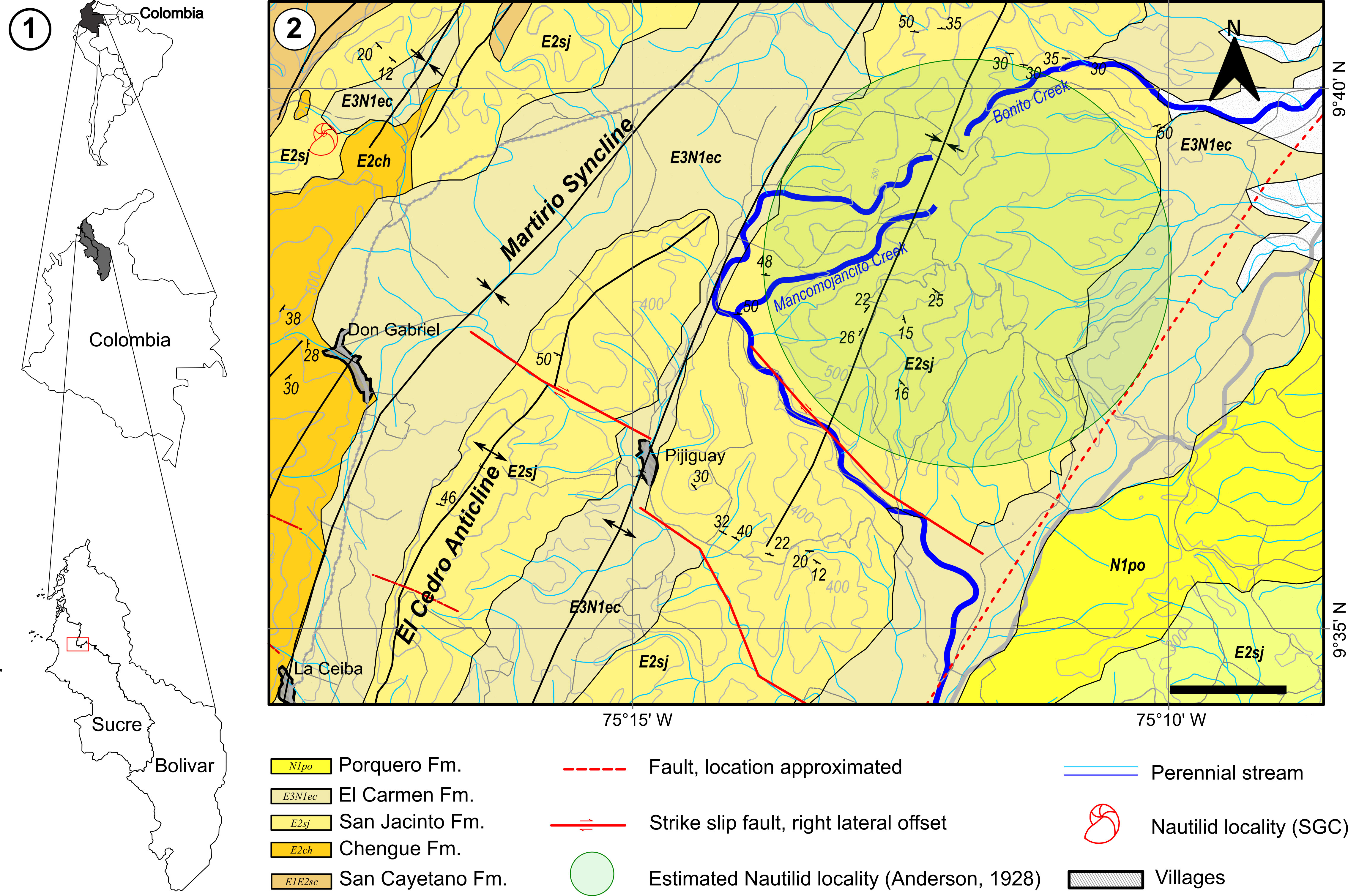
The 13 nautilid specimens were found in the Montes de María region, on the western margin of the San Jerónimo mountain range (Duque-Caro, 1979), on the southeastern termination of the Piedra de Moler Syncline, near the small town of Chengue (Sucre). The nautilids were recovered from a fossiliferous level exposed along a bedding plane near an intersectional road connecting Chengue and the Orejero region. This level is notably rich in fossils, most of which can be directly observed and collected due to the weathering of the calcareous material.
The nautilid specimens were found in a medium to thick bed of grainstone biosparite, with trace fossils of Thalassinoides Ehrenberg (1944), mollusks (gastropods and bivalves), rhodoliths, lepidocyclinines, Nummulites Lamarck (1804), arborescent corals, and bryozoans. The nautilids are preserved as internal molds of phragmocones with dimensions ranging between 10 and 25 cm in diameter. This fossiliferous level is located near the top of the San Jacinto Formation.
Overlying this fossiliferous level, there are conglomerates with bioclasts and small to medium-sized pebbles of subangular milky quartz and black and brown chert. The bioclasts are mostly 1–2 cm disarticulated bivalves and a minority of gastropods. Articulated bivalves with angular ribs 2 mm high are also found, although in smaller numbers. Pebbles are discoidal and subangular, and a small percentage of fragments are volcanic.
The San Jacinto Formation, from which the A. peruviana specimens reported here were recovered, has a stratigraphic range spanning the upper Eocene to the lowermost Oligocene according to Duque-Caro et al. (1996). This range assignment is further supported by Guzmán Ospitia et al. (2004), who proposed a slightly broader upper Eocene to lower Oligocene range. Biostratigraphic constraints for this formation are based on planktonic foraminiferal biozones P.15 to P.18 and the benthic biozone of Bulimina jacksonensis Cushman (1925), as documented by Guzmán Ospitia et al. (2004). Furthermore, Guzmán Ospitia (2007) supports a range from the upper Eocene to lower Oligocene based on indirect stratigraphic correlation.
As the specimens were collected during a cartography project, no stratigraphic section is available from the locality to ascertain a more precise position of the fossiliferous level. Therefore, the known stratigraphic range of the studied specimens would correspond to the one for the San Jacinto Formation, upper Eocene to lower Oligocene.
Following the proposals of Flügel (2010), we interpret that the upper part of the San Jacinto Formation was deposited in a subtidal environment setting on an open to restricted inner ramp, characterized by moderate to high energy conditions and the development of shallow sandbanks and reefs located above the wave base level. Anderson (1928, pp. 5–6), when referring to the locality of the nautilid finding, wrote: “Six miles southwest of El Carmen, on the west flank of the high anticlinal ridge”. Following this description, we located the site between the Mancomojancito and Bonito creeks, on the western flank of the San Jacinto Anticline (Angarita et al. (1983); Fig. 1) within the San Jacinto Formation (Duque-Caro et al., 1996). Based on this information, we estimate that the nautilid reported by Anderson (1928) was found approximately 13.5 km from the Chengue – Orejero region, where the 13 specimens presented in this contribution were found. Furthermore, we interpret that the nautilid specimens reported by Anderson (1928), along with the 13 new specimens, originate from the same stratigraphic unit: the upper part of the San Jacinto Formation.
2 MATERIALS AND METHODS
We describe 13 nautilid specimens (WC185-1, WC185-3–10, WC185-12–15), which are housed in the collections of the Museo Geológico Nacional José Royo y Gómez of the Servicio Geológico Colombiano (Bogotá, Colombia) under the acronym WC.
Measurements were taken from the best-preserved specimens (WC185-1, WC185-3, WC185-4, WC185-6, WC185-7, and WC185-12) using a vernier caliper. The following parameters were measured: diameter (D), whorl height (Wh), whorl width (Ww), and umbilical diameter (U). Additionally, in some cases we measured the central height (Ch), ventral distance (Vd), siphuncle diameter (Sd), and siphuncle index (Si=Vd/Ch) following Cichowolski et al. (2005) and Pohle et al. (2022). Morphometric ratios were calculated from these linear measurements to characterize the conch proportions (Tab. 1). The suture line of the specimen WC185-6 was drawn using transparent tape, digitized and subsequently edited. For the systematic assessment, we principally followed Kummel et al. (1964).
The suprageneric classification of the genus Aturia has changed over time. Kummel (1956), following d’Orbigny (1840), initially grouped all post-Triassic nautilids, including the subfamily Aturinae, within the family Nautilidae. He recognized Aturia as the sole genus within this subfamily, characterized by its dorsally positioned siphuncle. However, Kummel et al. (1964) later revised this classification, accepting the separate family Aturiidae as proposed by Chapman (1857). They argued that the greater complexity of the suture line in Aturia justified its familial distinction from the Nautilidae. We followed this latter proposal, as the suture line of Aturiidae exhibits a ventral saddle, a deep lateral lobe, and a lateral saddle, conforming to the morphological description provided by Hyatt (1894).
3 SYSTEMATIC PALEONTOLOGY
Class CEPHALOPODA Cuvier, 1797
Subclass NAUTILOIDEA Agassiz, 1847
Order NAUTILIDA Agassiz, 1847
Suborder NAUTILINA Agassiz, 1487
Family ATURIIDAE Chapman, 1857
Genus Aturia Bronn, 1838
Type species. Nautilus aturi Basterot, 1825. Cenozoic, Dax, Francia.
Aturia peruviana Olsson, 1928
Figures 2–5
1928 Aturia alabamensis var. peruviana n. var. Olsson, p. 60, pl. 24, figs. 1–3.
1928 Aturia colombiana n. sp. Anderson, p. 17, figs. 2–3, pl. 1, fig. 10.
1937 Aturia peruviana Olsson, Miller & Thompson, p. 64, pl. VIII, figs. 1–4, pl. IX, fig. 5
1939 Aturia peruviana Olsson, Miller & Furnish, p. 77, pl. 13, figs. 1–2, pl. 14, fig. 2
1946 Aturia colombiana Anderson, Clark in Clark & Durham, p. 12, pl. 1, fig. 1
1946 Aturia alabamensis var. peruviana Olsson, Clark in Clark & Durham, p. 12, pl. 1, figs. 2–3
1947 Aturia peruviana Olsson, Miller, fig. 21, fig. 25A, p. 102, fig. 28, pl. 12, fig. 2, pl. 78, fig. 1, pl. 79, figs. 1–2, pl. 80, figs. 1–2, pl. 81, figs. 1–4, pl. 82, figs. 1–3, pl. 83, figs. 1–3, pl. 84, figs. 3–4, pl. 85, figs. 1–3, pl. 86, figs. 1–2.
2012 Aturia peruviana Klug & Etter, fig. 4.12
Diagnosis. See Olsson (1928).
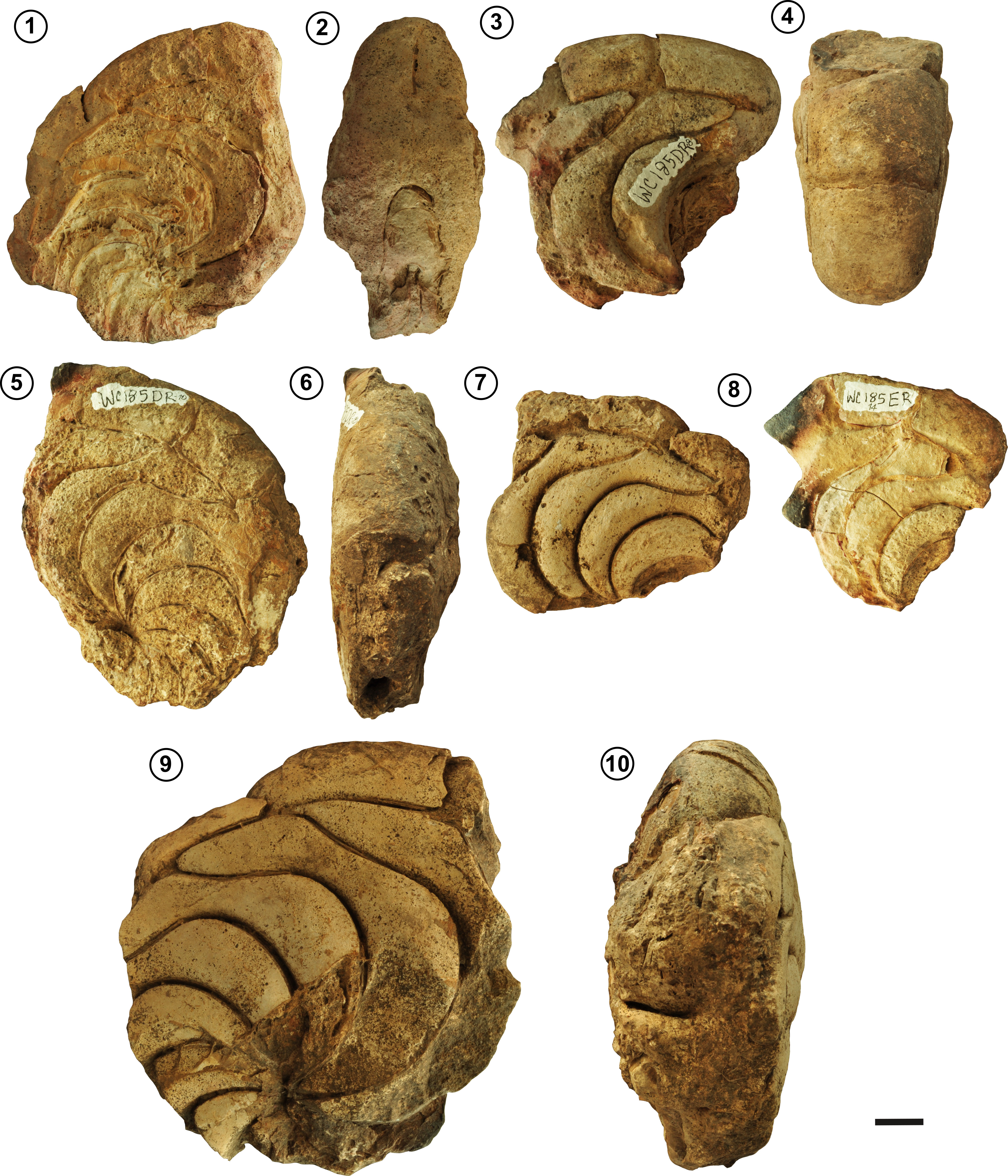
Referred material. 13 internal molds of phragmocones (WC185-1, WC185-3–10, WC185-12–15).
Table 1: Measurements (mm) and proportions of some specimens of Aturia peruviana.
| Specimen | D | Wh | Ww | U | Ch | Vd | Sd | Si | U/D | Wh/D | Ww/Wh |
|---|---|---|---|---|---|---|---|---|---|---|---|
| WC185-1 | - | 140 | 110? | - | ~148.3 | 91 | 26 | ~0.613 | - | - | 0.785? |
| WC185-3 | - | ~190 | - | - | ~128 | 94.6 | 24 | ~0.73 | - | - | - |
| WC185-4 | - | - | - | - | 135 | 88.6 | 25.5 | 0.656 | - | - | - |
| WC185-6 | ~221 | ~135.3 | 92.7 | 5 | 114.4 | 74.4 | - | 0.65 | ~0.022 | ~0.612 | ~0.636 |
| WC185-7 | - | 146 | 112 | - | - | - | - | - | - | - | 0.767 |
| WC185-12 | - | 142 | 97? | - | 129.8 | 81.1 | 33.7 | 0.624 | - | - | 0.683? |
Abbreviations: Ch, central height; D, diameter; Vd, ventral distance; Sd, siphuncle diameter; Si, siphuncle index (Vd/Ch) following Cichowolski et al. (2005); U, umbilical diameter; Wh, whorl height; Ww, whorl width.
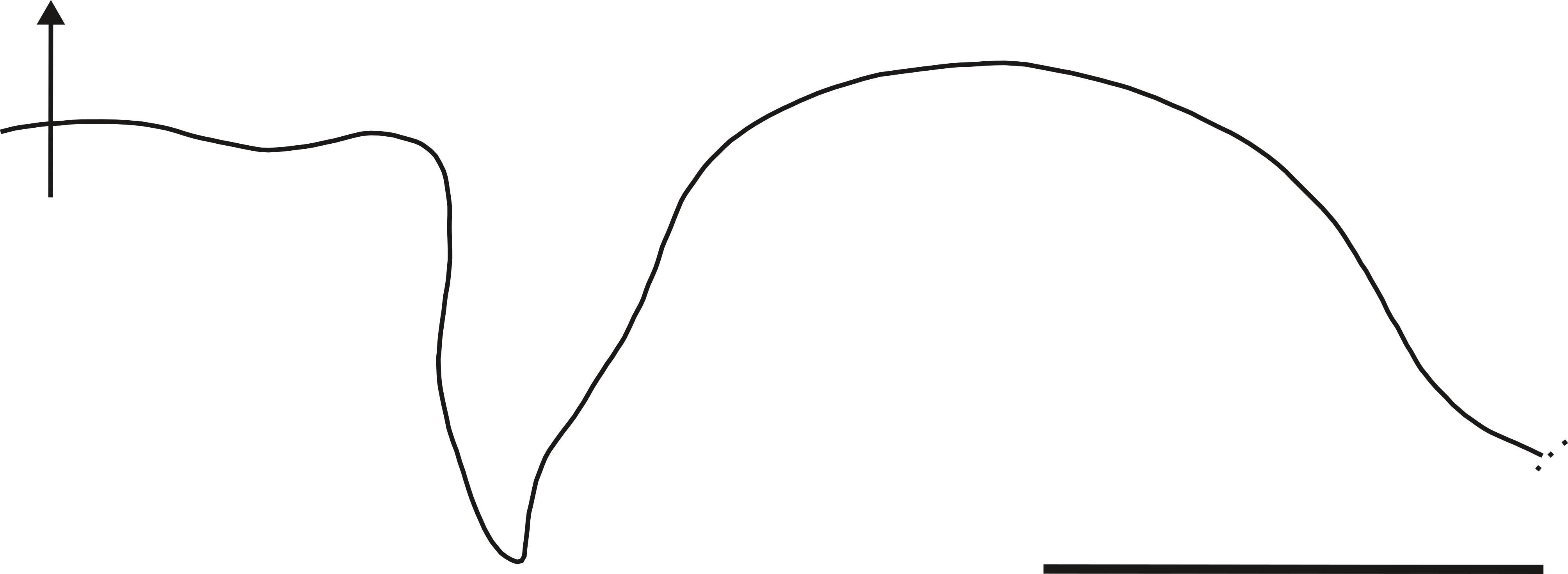
Description. Large adult phragmocones, reaching up to 22 cm in diameter. The conch is nautiloconic and involute (U/D = ~2.26%), with a compressed suboval whorl section (Ww/Wh = 66.3 to 78.5%). The flanks are predominantly arched, and the venter is rounded (Figs. 2.2–3, 2.5; 3.2–4, 3.8; 4.2, 4.4, 4.6, 4.10). The suture line features a broad, slightly flattened ventral saddle, a broad and very shallow ventrolateral lobe, and a somewhat narrow, slightly sinuous, and deep asymmetrical lateral lobe with a sharp lower contour, followed by a broad arched lateral saddle (Figs. 2.1, 2.4; 3.1, 3.5, 3.7; 4.3, 4.5, 4.7–9; 5). The siphuncle is relatively large and located between the center and the dorsum (average Si= 0.65; (Figs. 2.3, 2.5; 3.2–3). When preserved, the shell wall is smooth and relatively thin (Fig. 2.2), consistent with the description of Aturia by Miller (1947).
Remarks. The siphuncle of A. peruviana is relatively large, with proportions similar to those of Cymatoceras perstriatum (Cichowolski, 2003). The shell wall is notably thin; for example, in specimen WC185-1, a wall thickness of 1.3 mm at a projected diameter of 286 mm gives a diameter-to-thickness ratio of approximately 220:1. In contrast, a specimen of Nautilus pompilius with a diameter of 79 mm has a wall thickness of approximately 1 mm (pers. obs. by PP), yielding a ratio of about 79:1. This marked difference indicates that the shell of A. peruviana is proportionally much thinner than that of N. pompilius.
Geographic and Stratigraphic Occurrence. Aturia peruviana has been reported in Peru (Miller, 1947; Olsson, 1928), Colombia (Miller, 1947; Miller & Furnish, 1939), Panama (Miller, 1947; Miller & Furnish, 1939), Venezuela (Klug & Etter, 2012; Miller, 1947), Curazao (Jung, 1974), and probably Ecuador (Miller, 1947). According to Olsson (1928), Aturia peruviana occurs in Peru from the upper Eocene to the Oligocene. Miller & Thompson (1937) extended this range from the upper Eocene to the upper Oligocene, while Miller (1947) reported the species in upper Eocene and Oligocene deposits of Peru. Nielsen et al. (2009) suggested a broader stratigraphic range for Aturia, from the Eocene to possibly the lower Pliocene.
4 DISCUSSION
4.1 Historical reports of Aturia colombiana and Aturia peruviana in South America.
The genus Aturia was first reported from the Cenozoic of northern Colombia with the species A. colombiana Anderson, 1928 (Anderson, 1928; Clark & Durham, 1946) and A. cubaensis (Lea, 1841; in Jung 1966). Schenck (1931), as cited in Miller & Thompson (1937), Miller & Furnish (1939) and Miller (1947), later proposed that A. colombiana was a junior synonym of A. peruviana Olsson, 1928. Anderson (1928) recognized A. colombiana as a new species from the Eocene. In the same year, Olsson (1928) reported the presence of A. peruviana from the Talara Formation of Peru, assigning it to the upper Eocene and establishing it as a variety of Aturia alabamensis Morton (1834); a species originally described from the Eocene of Alabama, and subsequently documented along the southeastern Atlantic coast of the USA (Miller, 1947). Additionally, Olsson (1928) proposed a synonymy between A. colombiana and A. peruviana; however, he questioned this conclusion due to the poor preservation of the specimens, which made it difficult to determine whether they belong to a single species. Clark & Durham (1946), referring to the works of Anderson (1928) and Olsson (1928), nevertheless, concluded that A. colombiana and A. peruviana were two different species.
The specimen reported by Anderson (1928) was illustrated and described by Clark and Durham (1946, pl. 1, fig. 1). A color photograph of the medium-sized individual reported by Anderson (1928) is available on the California Academy of Sciences website https://researcharchive.calacademy.org/research/izg/fossil/Index.asp, showing a suture line with the same path between the lateral saddle and lateral lobe of A. peruviana. Klug and Etter (2012, Figure 4.12) also showed an internal mold of A. peruviana from Venezuela.
Taxonomic considerations. Jung (1966) assigned a Miocene specimen from the La Guajira Peninsula in Colombia, near the Venezuelan border, to A. cubaensis (a species that has also been reported from Venezuela) (Jung, 1966; Klug & Etter, 2012, 2013). The suture line of A. cubaensis, as described by Miller (1947), has a broad ventral saddle with a very shallow ventrolateral lobe, a deep and relatively wide lateral lobe, with a sharp rounded horned termination, and a highly arched and rounded lateral saddle (cf. Miller & Downs, 1950, pl. 9, fig. 3–4). The lateral lobe of A. cubaensis differs from that of A. peruviana in the outline as it has more pronounced curvatures and a narrower termination (cf. Anderson, 1928, pl. 1, fig. 10; Clark & Durham 1946, pl. 1, fig. 1; Miller, 1947, Figure 28B,).
The lateral lobe of A. peruviana shows a narrower termination and an outline with a variation in the curvature between the lobe and the lateral saddle (Fig. 5, cf. Miller & Thompson, 1937, Figure 2A) whereas the outline of A. cubaensis does not show this change in the saddle arch, which is imperceptibly connected with the lobe. The width and height of the lateral saddle of A. cubaensis are very different from those of A. peruviana (cf. Miller, 1947, pl. 83, figs. 4-6). The suture line of A. peruviana is similar to that of A. panamensis (cf. Miller, 1947, pl. 80, fig. 4) and to that of A. alabamensis (Miller, 1947, Figure 7F). According to Miller (1947) the sutures of A. alabamensis and A. peruviana are closely similar, however, these species can be differentiated based on the width of their conch. Nevertheless, Miller (1947) does not discard that these differences are due to sexual dimorphism.
5 CONCLUSIONS
The 13 newly described nautilid specimens are assigned to A. peruviana. These specimens originated from the San Jacinto Formation, consistent with the reported locality of nautilids documented by Anderson (1928). Morphological characteristics of the phragmocone and the suture line of the new material supports the synonymy of A. colombiana with A. peruviana, thereby corroborating the taxonomic revision proposed by Miller (1947). Furthermore, these findings indicate that the stratigraphic range of A. peruviana in Colombia is upper Eocene to lower Oligocene.
6 ACKNOWLEDGEMENTS
This work was supported by the Servicio Geológico Colombiano, project no. 1000239 “Operación para trampas de hidrocarburos asociados a los diapiros” completed in 2019, which enabled the field campaigns that resulted in the collection of the fossil specimens presented in this study. We are grateful to the editor, Marcela Cichowolski (Universidad de Buenos Aires, Argentina), and the reviewers, Ninon Allaire (CICTERRA, Córdoba, Argentina) and Marcin Machalsky (Polish Academy of Sciences, Poland), for their constructive feedback on our manuscript; their insights significantly enhanced the final version.
